第十三章
進化論的主張與事實
在前面的章節裏,我們依據化石記錄和分子生物學所提供的證據,分析了進化論的無效性。在本章中,我們將闡述進化論者作為理論根據的一些生物學現象和概念。這些話題尤為重要,因為它們顯示:並沒有支持進化論的任何科學發現,而且把進化論者的曲解與蒙蔽公佈於眾。
變異與物種
“變異”是遺傳學的專有名詞,涉及遺傳方面的事件;它引起某一物種的個體或群體具有互不相同的特徵。例如,地球上所有的人類,基本上擁有相同的遺傳信息,但有些人則有所不同,如斜眼、紅發、長鼻,或者有人體形矮小。這取決於遺傳信息潛在的變異。
進化論者把一個物種內部的變異,當作其理論的依據。然而,變異並不構成進化論的依據;因為,變異只是已經存在的遺傳信息,不同結合的結果,它們沒有給遺傳信息添加任何新的特性。可是,對進化論至關重要的問題是,嶄新的資訊如何造出嶄新的物種。
變異總在遺傳信息的限度內發生。遺傳學把這個限度稱為“基因池”。一個物種在基因庫中的所有特性,由於變異而出現各種各樣的方式。例如,作為變異的結果,在某些爬行動物中,相對地出現較長的尾巴或較短的腿子;因為長腿和短腿的資訊,在這些動物的基因庫中同時存在。但是,變異不會通過給爬行動物增加翅膀或羽毛,或者改變其新陳代謝的方式,使它們轉變成鳥類。這樣的變化,需要增加生物的遺傳信息;這種情形在變異中絕不可能存在。
達爾文在闡述他的理論時,並未意識到這個事實。他認為變異毫無限度。1844年,他在一篇文章中寫到:“大多數作者都假定,自然界的變異的確存在限度,雖然我無法揭示作為這個信念基礎的哪怕一個事實。”151在《物種起源》一書中,他把不同的變異例子,作為其理論的最大證據。例如,根據達爾文的觀點,為了獲得新品種的高產奶牛,動物飼養者讓奶牛與不同的種牛交配,最終把牛轉變成另一不同的生物。《物種起源》一書中的幾句話,最能說明達爾文“無限變異”的觀念:
“我能夠看得出,熊類通過自然選擇,獲得更多適應于水中生存的習性,形成愈來愈大的嘴巴,直到成為鯨魚那樣大的動物。”152
熊是從鯨魚進化來的嗎?

達爾文在《物種起源》中聲稱,鯨魚從試圖游泳的熊進化而來!達爾文錯誤地假定:在某一物種內,變異的可能性毫無限制。20世紀的科學已經表明,進化論純粹是想像出來的假想。
達爾文引用如此牽強的例子,是因為那個時代的人們,對科學原始的理解程度。那以後,20世紀的科學,基於生物實驗而提出了“遺傳穩定性”(遺傳的動態平衡)的原理。該原理認為,通過配種試圖產生新的變異,並不是決定性的手段,而不同生物之間有著不可逾越的屏障。這意味著動物飼養員,絕不可能通過不同動物之間的交配,把牛轉變成像達爾文假定的另一不同的物種。
駁斥進化論的諾曼•麥克佩斯在他的《重審達爾文》一書中寫到:
“問題的核心在於,生物是否確實毫無限度地改變形態……。物種是穩定的。我們已聽到了飼養員們的失望;他們沒能推進將動植物回歸到它們起點的工作。儘管經過兩三個世紀的不斷努力,但他們從來沒有生產出蘭玫瑰或黑鬱金香。”153
盧瑟•波班克是最有名的動物飼養專家,他針對這方面的事實說:“可能的發育存在著限度,並且這些限度遵循規律。”154 丹麥科學家W.L.詹森就此評論說:
“達爾文和華萊士強調的變異,不可能有選擇地推到某一點上,即這樣的可變性並不包括模糊起點中的秘密。”155
抗生素的抵抗力和DDT的免疫性不是進化論的證據
進化論者當作證據,努力拿出來的一個生物概念,是細菌對抗生素的抵抗力。幾乎所有的進化論資料顯示,他們把這種抗生素的抵抗性作為“生物通過有利突變而發育的例子”。類似的主張同樣,應用於昆蟲殺蟲劑的免疫性機制,如DDT(二氯二苯三氯乙)。
然而,進化論者在這個問題上同樣錯了。
抗生素是從微生物中產生的、抗擊其他微生物的“分子殺手”。最早的抗生素是青黴素(盤尼西林),由亞歷山大•弗雷明(fleming)於1928發現。弗雷明認識到,黴菌中產生可以殺滅葡萄狀球菌的分子,而這一發現是醫學界的一個轉捩點。抗生素來自抗細菌的微生物,其結果是成功的。
不久,發現了一些新東西。細菌隨時間的過去,形成了對抗生素的抵抗力。該機制是這樣運作的:大量的細菌造成抗生素的死亡,而另一些不受抗生素影響的細菌,快速繁殖並組成了整個細菌隊伍。這樣,所有的細菌對抗生素產生抵抗。 進化論者試圖把這種現象看成“細菌適應環境的進化”。
但是,真理遠遠不同於這種膚淺的解釋。在這方面做過詳細研究的以色列生物物理學家、因1997年出版《並非偶然》一書而聞名的李•斯普特納(Lee Spetner),主張細菌的抵抗性來自兩種不同的機制,但沒有一種會成為進化論的證據。這兩種機制是:
1, 轉變抵抗力的基因已經在細菌中存在;
2, 細菌形成對抗生素的抵抗,是由於突變而失去其基因資料的結果。
斯普特納教授在2001年發表的一篇文章中,解釋了第一種機制:
“一些微生物具有抵抗這些抗生素的基因。這種抵抗以退化抗生素分子或將之逐出細胞的形式實現……。具備這些基因的機制,會把它們轉移給其他細菌,使之也產生抵抗力。雖然抵抗機制對某種抗生素有效,但大多數病原體細菌,已成功地積累幾組基因,由它們對不同的抗生素產生抵抗力。”156
於是,斯普特納進一步說,這不是“進化的證據”:
“以這樣的方式,產生對抗生素的抵抗力……不是用來說明進化論突變之說的原型。可能注解該理論的遺傳性變化,不必只是給細菌的基因組增加資訊,而必須給biocosm增加新資訊。基因水平式的轉變,僅僅分散一些生物中已存在的基因。”157
所以,我們在此不能談論進化論,因為沒有產生新的遺傳信息:已存在的遺傳信息只在細菌間轉化。
由突變產生的第二種機制,也不是進化的例子。斯普特那寫道:
“……有時候,微生物通過單一核苷的取代作用,獲得對抗生素的抵抗力……。鏈黴素由賽裏曼•沃克茨曼(Selman Waksman)和阿爾伯特•斯凱茨(Albert Schatz)發現,並於1944年首次報告;它是細菌用這種方式獲得抵抗力的一種抗生素。可是,儘管他們經歷的突變,對鏈黴素面前的微生物有益,可它不能當作新達爾文理論(NDT)的那種突變所需要的原型。抵抗鏈黴素的那種突變,在核糖體中是明顯的,並使對抗抗生素分子的分子退化。這種在微生物核糖體表面上的變化,妨礙鏈黴素分子的抗生素作用。這一退化是鏈黴素作用的損失,因此,也是資訊的損失。關鍵在於,不論有多少抵抗力,進化不可能從這種突變中獲得。進化不能建立在僅僅積累使原有特性退化的突變上。”158
概括而言,與細菌核糖體接觸的突變,使細菌抵抗鏈黴素。其原因是突變“分解”了核糖體。也就是說,細菌中沒有增加新的遺傳信息。相反,核糖體的結構被分解了,即細菌“殘廢”了。(還有,研究發現,變異細菌的核糖體的作用,不如正常細菌的作用。)由於這種“無能”妨礙使之附著於核糖體的抗生素上,所以“抗生素抵抗力”發育出來了。
最後,並沒有出現“發育遺傳信息”突變的例子。
同樣,昆蟲對DDT及類似殺蟲劑的免藥力也確有其事。大多數例子說明,已經存在的免疫基因都被利用了。進化論生物學家弗蘭西斯克•阿亞拉承認這個事實:“遺傳的量變對大多數殺蟲劑有抵抗力;這在現有的每一種人造混合物中明顯地暴露了。”159 突變解釋的其他例子,就像上面提到的核糖體的突變,是導致昆蟲“遺傳信息不足”的現象。
在這種情況下,誰都不能聲稱,細菌和昆蟲的免疫機制,是構成進化理論的證據。那是因為,進化論主張生物通過突變而發育。但是,斯普特納解釋說,抗生素的抵抗力和別的生物現象,都沒有顯示過如此突變的例子:
“從未有人觀察到需要種外進化(大進化)的突變。表現新達爾文理論需要的那種隨意的突變,就在分子階段一無所獲地完蛋了。我的問題是:用來支撐這種理論的突變,還需要支援嗎?回答只能是‘不!’”160
退化器官的謬論
很久以來,“退化器官”的概念作為進化的“證據”頻繁出現在進化論文獻中。當證明該概念無效後,它終於悄然擱置起來了。但是,一些進化論者仍然相信它,並且有人不時努力把它作為進化論重要的證據。
“退化器官”的觀念,最早在一個世紀以前提出。像進化論宣稱的那樣,某些生物的體內存在一些非功能性的器官,是從祖先那裏繼承的、不被使用而逐漸退化了的器官。這個假定很不科學,缺乏必要的知識。這些“非功能性器官”,實際上是“功能還沒有被發現”的器官。
最能說明這種提法的,是進化論者所列舉的長長的“退化器官表”。進化論者S.R.斯卡丁(Scadding)在《進化理論》(Evolutionary Theory)雜誌發表“退化器官能構成進化論的證據嗎?”一文,談到了這個事實:
“既然不可能含糊地鑒別無用的器官,用來爭論的器官沒有科學依據,那我斷定:‘退化器官’給進化論提不出任何特別的證據。”161
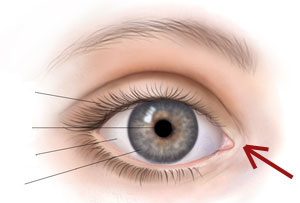
所有“退化器官”的例子隨時日而被否定。如,達爾文在《物種起源》中說成“退化器官”的眼睛中的淚器,現在已證實完全有作用;雖然它在達爾文的時代尚不知道。這個器官清潔和潤滑眼球。
德國解剖學家威德希姆(R.Wiedersheim)1895年列出的退化器官清單,包括闌尾、尾骨等大約100個的器官。隨著科學的發展,人們發現威德希姆列表中的所有器官,其實在人體內有非常重要的作用。例如,被當成“退化器官”的闌尾,實際上是與人體內的感染作鬥爭的淋巴器官。1997年,這個事實清楚了:“身體的其他器官與組織:胸腺、肝、脾臟、闌尾、骨髓,以及一些小的淋巴組織,如喉內的扁桃體和小腸中的皮爾補丁(Peyer's patch)等,也是淋巴系統的部分。它們同樣幫助身體與病菌作戰。”162
列入“退化器官”中的扁桃腺,在保護喉嚨抗感染方面,扮演著重要角色,尤其在人的青春期;位於脊椎骨末端的尾骨,起著支撐盆骨和收縮肌肉的作用;因此,沒有尾骨就不可能坐得舒服。在隨後的幾年中,人們認識到,胸腺借助T細胞啟動人體內的免疫系統;松果腺負責一些重要激素的分泌;甲狀腺作用于嬰兒和孩子的穩定成長;腦垂體控制許多激素腺的正確運作。而所有這些器官曾被當作“退化器官”。後來,達爾文看成“退化器官”的眼睛中的淚器,實際上負責清潔和潤滑眼球。
進化論者關於退化器官的斷言,有一個非常重要的邏輯錯誤。正如我們剛才解釋的那樣,進化論者聲稱生物的退化器官,是從它們的祖先那裏繼承的。然而,一些所謂“退化了的”器官,並沒有在稱為人類祖先的生物中發現過!例如,闌尾在被認為是“人類祖先”的猿類中並不存在。挑戰“退化器官”理論的著名生物學家H.伊諾克,論述了這個邏輯錯誤:
“猿擁有闌尾;由於沒有直接的親緣關係,較為低級的猿卻沒有闌尾;袋鼠也有闌尾。進化論怎能解釋這些現象呢?”163
進化論者匆匆提出的退化器官的假想,存在許多邏輯缺陷,並犯了科學上的錯誤。人體根本不存在所謂從祖先那裏繼承的任何退化器官;因為,人類並非從其他生物偶然地進化而來,而是以現有、完整、俊美形式的創造。
“異體同形”的神話
在生物學中,把不同物種之間結構上的相像叫作“異體同形”。進化論者試圖將那些相像作為進化論的證據。
達爾文認為器官(同族體)相像的動物,互相有進化的關係,而這些器官一定是從一個共同祖先那裏繼承的。按照他們的假定,鴿子和鷹都有翅膀,這樣,它們和全部有翅膀的其他鳥類,是從一個共同的祖先進化而來的。
異體同形是毫無依據的騙人的論點,它只是憑外觀形體上的相像而提出的假設。自達爾文時代起,至今沒有任何具體的發現,來證實這個論點。任何地方也沒有發現這一想像中的“異體同形”之共同祖先的化石。而且,下列論點清楚表明,同族體並沒有提供發生進化的證據:
1,人們發現同族體的器官屬於不同的動物物種,進化論者在它們當中,沒能建立任何一種進化關係;
2,某些具有同族體器官之生物的遺傳密碼,完全互不相同;
3,不同生物同族體器官的胚胎發育階段完全不同。
現在我們對這些要點逐一加以分析。
完全不同生物的相像器官
在進化論者無法建立任何進化關係的不同物種間,共用不少相似的(同源性)器官。翅膀就是其中的一個例子。除了鳥類,作為哺乳動物的蝙蝠也有翅膀,並且昆蟲甚至一些滅絕的恐龍也有翅膀。進化論者無法在這四種不同種類的動物中,建立任何進化關係或親緣關係。
另一個明顯的例子,是不同生物的眼睛驚人的相似和結構上的類似。例如,章魚和人是兩種無法建立任何進化關係的不同生物,而兩者的眼睛在結構與功能上卻極為相似。就算他們的眼睛很相似,進化論者也不能聲稱人和章魚具有共同祖先。這些以及許多別的例子證實,進化論者所謂“同源性器官證明生物從一個共同祖先進化而來”的主張並無科學依據。
實際上,同源性器官使進化論者相當尷尬。著名進化論者弗蘭克•索利斯伯裏(Frank Salisbury)在談到不同物種具有非常相似的眼睛時說:
“像眼睛一樣複雜的器官,在其他群體中出現過,如魷魚、脊椎動物和節肢動物。把這些生物的起源解釋上一次,事情就夠糟糕的了;但根據現代合成理論(新達爾文學)的觀點,把它們想上幾次,則使我一頭霧水。”164
儘管許多生物的形體極為相似,但這並不支援它們之間有進化關係的主張。兩種大型哺乳動物,如胎生動物和有袋動物就是例子。進化論者承認,這兩種哺乳動物一開始就彼此分開,並完全獨立地發育。然而,在這兩類生物中,形體非常相似的大量動物,據說已徹底滅絕了。美國生物學家丁•肯揚和潑斯沃爾•大衛斯(Dean Kenyon and Percival Davis)評論道:
“根據達爾文理論,狼、貓、松鼠、地豬、食蟻動物、鼴鼠和老鼠需要進化兩次:第一次進化成胎生動物,第二次完全獨立地進化成有袋動物。這等於隨意、不受指導的突變和自然選擇的驚人主張:在廣為分散的生物體中,偶然地找到同樣的特徵。”165
像這些極其接近和類似的器官顯示,從共同祖先進化的理論並沒有證據;進化論生物學家不能把“同源體”作為其理論的例子。
異體同形在遺傳與胚胎學上的僵局
為了認真對待進化論者“異體同形”的主張,不同生物的類似(同源性)器官,應該用類似的DNA密碼編碼。但是,事情並非如此。在多數情況下,遺傳密碼是完全不同的。而且,不同生物在DNA中相似的遺傳密碼,常常與不同的器官相聯。
澳大利亞生物化學教授邁克爾•登頓在其《進化論:危機中的理論》(Evolution:A Theory in Crisis)一書中,描述了進化論者對異體同形,在遺傳學上的解釋所面臨的尷尬:“異體同形結構常常列入非同源體遺傳系統中,而同源體的概念很少擴大到胚胎學的範圍中。”166
這方面的著名例子,是幾乎在進化論所有教科書中引用的四足動物的“五位元數骨骼結構”。四足動物即生活在陸地上的脊椎動物,在前後爪子上的指關節都是五位數。儘管它們並不像我們知道的那樣,但由於它們的骨頭而被認為長有五指。青蛙、蜥蜴、松鼠或猴子的前後爪也是那樣。鳥類和蝙蝠的骨骼也有這種基本的結構。
進化論者關於所有生物來自一個共同祖先的主張,其長期引用的證據就是“五指”。這一主張是20世紀的生物學,把它作為進化論有力證據的基本源泉。但20世紀80年代的遺傳學發現,駁倒了進化論的這一主張。人們認識到,不同生物的“五指”,是由不同的基因控制的。進化論生物學家威廉姆•菲克斯,用這樣的方式描述了這一理論的破滅:
“進化論老式的教科書,大量利用了同源體進化的概念,即不同動物間肢體骨骼上的明顯類似。這樣,人身上的‘五指’型肢體、鳥的翅膀、鯨的鰭狀肢,都預示著它們共同的起源。如果這些形式多樣的結構,是由同樣的基因、由突變時而不同,並以自然選擇的形式遺傳的,那麽,該理論就有意義了。不幸的是,情況並非如此。人們現在知道,同源體器官完全是在不同生物中的基因產生的。從同一祖先傳承類似基因的同源體概念已經破產了……”167
還有一點,為了慎重地看待進化論的同源性,在相似結構的胚胎發育階段,即在卵階段或母體子宮內的發育時期,需要類似的發育;雖然在現實中,每一種生物在類似的胚胎階段彼此很不一樣。
結論:我們可以說,遺傳與胚胎方面的研究已證明,達爾文作為“生物從同一祖先進化的證據”的同源性概念,絕對不是證據。科學在這方面,一再證明了達爾文論題的錯誤。
分子同源性主張的無效性
進化論者把同源體作為進化的證據,不僅在形態學上無效,而且在分子上也是無效的。進化論者說,不同生物的DNA密碼或蛋白質結構是相似的,而這種相似是這些生物來自共同祖先或彼此進化的證據。
但真相在於,分子比較的結果對進化論根本無用。在看上去很相似的生物中,它們的分子有巨大的差異。例如,對呼吸至關重要的細胞色素C,在同類生物中具有難以置信的差異。根據這方面的研究結果,兩種不同爬蟲間的差異,要比鳥跟魚或魚跟哺乳動物的大。168 另一研究顯示,同一鳥類分子間的差異,比這類鳥與哺乳動物的還要大。人們還發現,看上去相似的細菌,在分子中的差異,要比哺乳動物、兩棲動物或昆蟲的大。在血色素、肌球素、激素和基因中也作了這樣的比較。169
考慮到類似發現,邁克爾•鄧肯評論道:
“在分子水平上的每一生物種類是獨特、孤立的,並且與任何媒介沒有關聯。因此,像化石一樣,分子沒有給長期尋找難懂的中間形式的進化論生物學家以證據……。在分子水平上,任何生物體沒有可以比作‘祖先’、‘原始’或‘高級’的親緣關係……。無疑,如果這些分子證據在一個世紀以前存在的話……器官進化的觀念也許根本不會存在。”170
20世紀90年代,對生物基因密碼的研究,加劇了進化論這方面進退維谷的尷尬。在實驗中,取代早些時候局限於對蛋白質序列的比較,而用“核糖體RNA”(rRNA)序列來比較了。從這些發現中,進化論科學家豎起了一棵“進化樹”。然而,這個結果使進化論者感到失望。根據法國生物學家赫維•菲力浦和帕垂克•福特拉(Hervé Philippe and Patrick Forterre)1999年寫的文章,“隨著越來越多的序列比較,大多數蛋白pyhlogenies彼此抵觸,rRNA樹也一樣。”171
除rRNA的比較外,生物基因中的DNA也得以比較。但是,結果與進化論者所預料的“生命樹”恰恰相反。1999年,分子生物學家詹姆士•萊克、拉維•傑因、馬萊亞•李沃拉在一篇文章中詳細描述:
“科學家對不同生物體的各種基因的分析發現,它們之間的關係與進化論的生命樹相衝突——僅僅源自對rRNA的分析。”172
組成蛋白質的比較和組成rRNA或基因的比較,都不是證實進化理論的前提。伊利諾斯大學有名的生物學家卡爾•沃塞承認,在分子發現的面前,生物的“發展史”概念失去了意義:
“從眾多已產生的個體蛋白的發育中,沒有出現生物體相容的發育歷史。動植物的不適應性,在普通樹上隨處可見,從根部到主幹,以及組成自身初級分組的各種(基)當中。”173
分子比較的結果,不是對進化論的贊同而是反對。1999年,《科學》雜誌上發表“到根除生命樹的時候了嗎?”一文,也承認這個事實。作者伊莉莎白•潘妮斯說,進化論生物學家對基因的分析與比較,本來想豎立“生命之樹”,但結果正好相反;並說“新資料正毀壞進化論的畫卷”:
“一年前,觀察從眾多微生物中重新排列的基因組的生物學家認為,這些資料可能支援生命早期的歷史。但是,他們對看到的一切卻迷惑了。當時對基因組的比較,不僅沒有澄清生命的主要元件進化的過程,而且對此大惑不解。現在,看著手裏掌握的另外八種微生物的序列,情況更加迷茫了……。許多進化論生物學家原以為,他們能夠大致看到生命開始的三個王國……當完整的DNA序列,打開與別的基因比較的道路時,研究者們卻期望,只是簡單地給這棵樹增加細節。但‘沒有什麽比真理更遙遠’,馬里蘭的‘基因組研究所’的負責人克雷爾•弗拉賽說。與研究者的初衷相反,比較產生了生命之樹的許多版本;它們不同於rRNA樹,而且互相衝突……。”174
概括而言,隨著生物學的發展,同源體概念失去了存在的土壤。由蛋白質、rRNA和基因組成的比較顯示,按照進化論的說法,那些近親的生物其實一一滅絕了。1996年的一次研究,使用了88種蛋白質序列,把兔子歸在靈長目而非嚙齒目動物中;1998年,分析了放在脊索動物海膽中的19種動物的13種基因;1998年基於12種蛋白質的另一次研究:使牛比馬更接近鯨。2000年,分子生物學家喬納森•維爾斯概括了這一情形:
“樹之間基於不同分子的矛盾,對這種怪誕之樹所作的分子分析,已使分子的發展歷史陷入危機。”175
胚胎重演的神話

在許多方面,赫克爾是個比達爾文更激進的進化論者。 因此,他毫不遲疑地歪曲科學資料,並作了各種各樣的偽造。
以前常說的“重述理論”早就從科學文獻中清除了,但仍有一些進化論的出版物,把它作為科學事實提出來。“重演”一詞是由進化論學生物學家厄恩斯特•赫克爾(Ernst Haeckel),在19世紀末提出的,是“個體發生學重演族系發展史”理論的概括形式。
這個理論假定,活胚胎再次經歷了其假冒祖先經歷的進化過程。赫克爾把在母體子宮裏的發育理論化了:人的胚胎首先顯示魚的特徵,然後是爬行動物的特徵,最後才是人的特徵。
據證實,這個理論完全是偽造的。人們現在知道,假定在人類胚胎早期出現的“鰓”,實際上是人的中耳導管、甲狀旁腺和胸腺的初級階段。 比作“卵黃囊”的胚胎部分,是為嬰兒生產血液的育兒袋。赫克爾和他的追隨者鑒定為“尾巴”那個部分,實際上是脊椎骨;它之所以像尾巴,只是因為它在腿子之前成型了。
在科學界,這是普遍公認的事實,甚至進化論者也接受這個事實。新達爾文主義的創始人之一喬治•蓋洛德•辛普森寫到:“赫克爾說錯了進化原理。現在可以肯定地確立,個體發生學並不重複族系發展史。”176
我們在《美國科學家》雜誌上讀到一篇文章:
“的確,生物起源的法則已經完全滅亡了,最終從20世紀50年代的生物教科書中除去了。作為一個受到質疑的嚴肅的理論話題,它在20年代就熄滅了……。”177
“重演”的另一有趣之處是,厄恩斯特•赫克爾為了支持自己提出的“重演理論”,成了偽造自己圖畫的高手。赫克爾的偽造企圖說明,魚和人的胚胎互為相似。當他露出馬腳後,唯一的答辯就是其他進化論者,已經承認犯過類似的錯誤:
海克尔伪造的图纸的

手,海克尔的胚胎的人类胚胎相似,为了证明鱼吸引了假照片。相对于真正的人类胚胎器官似乎已经被故意排除了大多数。 (Francis Hitching, The Neck of the Giraffe: Where Darwin Went Wrong", s. 205)
“對‘偽造’做出折衷的供認之後,如果看不到與我並肩工作的、數百位囚徒同事的安慰,我應該考慮自己受到譴責與消滅;那些同事中,有許多最值得信賴的觀察者和值得尊重的生物學家。他們出版的最好的生物學教科書裏,其中的大多數圖表、論文和雜誌,將受到與我的‘贗品’一樣的指控;因為,它們都是不準確的知識,或多或少地被篡改、公式化及編造了。”178
的確,有“數百位最值得信賴的觀察者和最值得尊重的生物學家”在搞研究;其中充滿帶著偏見的結論、曲解甚至編造。因為,他們總是習慣于維護進化理論,雖然沒有任何科學證據來支持這個理論。
笔记
151 Loren C. Eiseley, The Immense Journey, Vintage Books, 1958, p. 186. ![]()
152 Charles Darwin, The Origin of Species: A Facsimile of the First Edition, Harvard University Press, 1964, p. 184. ![]()
153 Norman Macbeth, Darwin Retried: An Appeal to Reason, Harvard Common Press, New York: 1971, p. 33. ![]()
154 Ibid, p. 36. ![]()
155 Loren Eiseley, The Immense Journey, Vintage Books, 1958. p. 227. ![]()
156 Dr. Lee Spetner, "Lee Spetner/Edward Max Dialogue: Continuing an exchange with Dr. Edward E. Max", 2001, http://www.trueorigin.org/spetner2.asp ![]()
157 Ibid. ![]()
158 Ibid. ![]()
159 Francisco J. Ayala, "The Mechanisms of Evolution", Scientific American, Vol. 239, September 1978, p. 64. ![]()
160 Dr. Lee Spetner, "Lee Spetner/Edward Max Dialogue: Continuing an exchange with Dr. Edward E. Max", 2001, http://www.trueorigin.org/spetner2.asp ![]()
161 S. R. Scadding, "Do 'Vestigial Organs' Provide Evidence for Evolution?", Evolutionary Theory, Vol 5, May 1981, p. 173. ![]()
162 The Merck Manual of Medical Information, Home edition, New Jersey: Merck & Co., Inc. The Merck Publishing Group, Rahway, 1997. ![]()
163 H. Enoch, Creation and Evolution, New York: 1966, p. 18-19. ![]()
164 Frank Salisbury, "Doubts About the Modern Synthetic Theory of Evolution", American Biology Teacher, September 1971, p. 338. ![]()
165 Dean Kenyon & Percival Davis, Of Pandas and People: The Central Question of Biological Origins (Dallas: Haughton Publishing, 1993), p. 33 ![]()
166 Michael Denton, Evolution: A Theory in Crisis. London, Burnett Books, 1985, p.145. ![]()
167 Fix, William, The Bone Peddlers: Selling Evolution (New York: Macmillan Publishing Co., 1984), p. 189 ![]()
168 W. R. Bird, The Origin of Species Revisited, Thomas Nelson Co., Nashville: 1991, pp. 98-99; Percival Davis, Dean Kenyon, Of Pandas and People, Haughton Publishing Co., 1990, pp. 35-38. ![]()
169 W. R. Bird, The Origin of Species Revisit-267 ed, pp. 98-99, 199-202. ![]()
170 Michael Denton. Evolution: A Theory in Crisis. London: Burnett Books, 1985, pp. 290-91. ![]()
171 Hervé Philippe and Patrick Forterre, "The Rooting of the Universal Tree of Life is Not Reliable", Journal of Molecular Evolution, vol 49, 1999, p. 510 ![]()
172 James Lake, Ravi Jain ve Maria Rivera, "Mix and Match in the Tree of Life", Science, vol. 283, 1999, p. 2027 ![]()
173 Carl Woese, "The Universel Ancestor", Proceedings of the National Academy of Sciences, USA, 95, (1998) p. 6854 ![]()
174 Ibid. ![]()
175 Jonathan Wells, Icons of Evolution, Regnery Publishing, 2000, p. 51 ![]()
176 G. G. Simpson, W. Beck, An Introduction to Biology, New York, Harcourt Brace and World, 1965, p. 241. ![]()
177 Keith S. Thompson, "Ontogeny and Phylogeny Recapitulated", American Scientist, Vol 76, May/June 1988, p. 273. ![]()
178 Francis Hitching, The Neck of the Giraffe: Where Darwin Went Wrong, New York: Ticknor and Fields 1982, p. 204. ![]()
- 特 別 前 言 达尔文主义与唯物主义 实为恐怖主义的理论根源
- 導 言 為什麽要談進化論?
- 序 言 相信進化論 是現代最大的奇跡
- 第一章 從偏見中解脫出來
- 第二章 進化論簡史
- 第三章 進化論想像中的機制
- 第四章 化石記錄反駁進化論
- 第 五 章 從水遷移到陸地的童話
- 第 六 章 鳥類與哺乳動物的起源
- 第 七 章 含欺詐成分的化石演繹
- 第 八 章 進化論的偽造品
- >第 九 章 人類進化的假想
- 第 十 章 進化論關於分子的僵局
- 第十一章 熱力學否定進化論
- 第十二章 設計與巧合
- 第十三章 進化論的主張與事實
- 第十四章 進化論的唯物主義傾向
- 第十五章 媒體:進化論的沃土
- >第十六章 結論:進化論是個騙局
- 第十七章 創造的事實
- 第十八章 物質的真正本質
- 第十九章 時間的相對論和命運的事實
- 第二十章 科學研究基金會(SRF)研 討會: 向公眾揭露進化論的活動