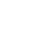
Kapitel 12: Varför evolutionisters hävdanden är ogiltiga.
Kapitel 12: Varför evolutionisters hävdanden är ogiltiga.
I tidigare kapitel, undersökte vi ogiltigheten i evolutionsteorin i form av bevis som finns i den fossila dokumentationen och med molekylär biologi som utgångspunkt. I detta kapitel kommer vi att behandla ett antal biologiska fenomen och begrepp som av evolutionister presenteras som teoretisk bevisning. Dessa frågor är särskilt viktiga eftersom de visar att det inte finns någon vetenskaplig slutsats som stödjer evolution men som istället avslöjar omfattningen av den distorsion och dupering som utnyttjas av evolutionister.
Variationer och arter.
Variation, en term som används inom genetiken, avser en genetisk händelse som ger upphov till olika egenskaper hos individen eller grupper av en viss typ eller art jämfört med varandra. Till exempel bär alla människor på jorden i stort sett samma genetiska information, men några har sneda ögon, några har rött hår, en del har långa näsor och andra är korta till växten, allt beroende på omfattningen av variationspotentialen i den genetiska informationen.
Evolutionister påtalar variationerna inom en art som bevis för teorin. Men variation utgör inte bevis för evolution, eftersom variationerna är resultaten av olika kombinationer av redan existerande genetisk information och de tillför ingen ny egenskap till den genetiska informationen. Det viktiga för evolutionsteorin, är dock frågan om hur helt ny information, för att göra en helt ny art, kan komma till stånd.
VARIATIONER INOM ARTER INNEBÄR INTE EVOLUTION
I boken The Origin of Species, förvirrade Darwin ihop två separata begrepp: variationer inom en art och uppkomsten av en helt ny art. Darwin observerade olikheter inom till exempel olika hundraser, och föreställde sig att en del av dessa variationer en dag skulle förvandlas till en annan art. Än idag fortsätter evolutionister att försöka beskriva variationer inom arter som "evolution".
Det är emellertid ett vetenskapligt faktum att variationer inom en art inte är evolution. Till exempel: oavsett hur många hundraser som finns, kommer dessa alltid att förbli en enda art - hund. Ingen övergång från en distinkt art till en annan kommer någonsin att äga rum.Variation sker alltid inom ramarna för genetisk information. I vetenskapen om genetik, kallas denna gräns "genpool". Alla de egenskaper som finns i genpoolen för en viss art kan uppdagas på olika sätt beroende på variationen. Till exempel, som en följd av variation, kan former som har relativt längre svans eller kortare ben förekomma i en viss art av reptil, eftersom information för både långbenta och kortbenta former kan finnas i genpoolen hos denna art. Däremot förvandlar inte variationer reptiler till fåglar genom att lägga till vingar eller fjädrar, eller genom att ändra deras metabolism. En sådan förändring kräver en ändring av den genetiska informationen, vilket sannerligen inte är möjligt genom variationer.
Darwin var inte medveten om detta faktum när han formulerade sin teori. Han trodde att variationer var gränslösa. I en artikel som han skrev år 1844 konstaterade han: "Att en gräns för variation existerar i naturen antas av de flesta författare, men jag kan inte upptäcka ett enda faktum på vilken denna tro är grundad”.143
Origin of Species citerade han olika exempel på varianter som de viktigaste bevisen för sin teori.
Till exempel, enligt Darwin, djuruppfödare som avlade fram avkomma från olika varianter av nötkreatur i syfte att åstadkomma nya varianter som producerade mer mjölk, skulle till slut omvandla dem till en annan art. Darwins uppfattning om "obegränsad variation" är bäst belyst i följande mening från The Origin of Species:
Jag ser inga svårigheter hos en ras av björnar som, genom naturligt urval, blir mer och mer vattenlevande i sina vanor, med större och större munnar, tills en varelse lika monstruös som en val producerades.144
Anledningen till att Darwin citerade ett långsökt exempel var den primitiva förståelsen för vetenskap på hans tid. Sedan dess, under 1900-talet, har vetenskapen antagit principen om "genetisk stabilitet" (genetisk homeostas), baserat på resultat av experiment som har utförts på levande ting. I denna princip gäller att eftersom alla parningsförsök som utförts för att avla fram nya varianter har varit resultatlösa, finns det strikta hinder mellan olika arter av levande varelser. Detta innebar att det var absolut omöjligt för uppfödare att omvandla nötkreatur till en annan art genom att para ihop olika varianter av dem, såsom Darwin hade antagit.
Norman Macbeth, som har motbevisat darwinism i sin bok Darwin Retried, säger:
MYTEN ATT VALAR UTVECKLADES FRÅN BJÖRNAR

I boken The Origin of Species, hävdade Darwin att valar hade utvecklats från björnar som försökte simma! Darwin antog felaktigt att möjligheterna till variation inom en art var obegränsade. 1900-talets vetenskap har visat att detta evolutionära scenario är inbillning.
”Kärnan i problemet är huruvida levande ting faktiskt varierar i en obegränsad utsträckning. … Arterna verkar stabila. Vi har alla hört talas om besvikna uppfödare som fört sitt arbete till en viss punkt bara för att se djur eller växter återgå till sina ursprungliga former. ”Trots stora ansträngningar under två till tre århundraden har det aldrig varit möjligt att producera en blå ros eller en svart tulpan”.145
Luther Burbank, ansedd som den mest kompetenta uppfödaren genom tiderna, uttryckte det när han sade: "det finns gränser för utvecklingens möjligheter, och dessa gränser följer en lag”.146
Den danska forskaren WL Johannsen sammanfattar saken så här:
Variationerna på vilka Darwin och Wallace hade lagt sin tyngdpunkt kan inte selektivt skjutas bortom en viss punkt, att en sådan variation inte innehåller hemligheten om "en obestämd ny linje”.147
På motsvarande sätt är de olika finkar som Darwin såg på Galapagosöarna ett annat exempel på variation som inte utgör bevis för "evolution". Observationer på senare tid har visat att finkar inte genomgår en obegränsad variation som Darwins teori förutsätter. De flesta av de olika typerna av finkar som Darwin trodde representerade 14 olika arter parade sig i praktiken med varandra, vilket innebär att de var variationer som tillhörde samma art. Vetenskaplig observation visar att finkars näbbar, som har varit mytomspunna i nästan alla evolutionistiska källor, i själva verket är ett exempel på "variation", och därför inte utgör bevis för evolutionsteorin. Till exempel har Peter och Rosemary Grant, som tillbringade åratal med att observera finkar på Galapagosöarna i sökande efter bevis för Darwinistisk evolution, tvingades att dra slutsatsen att ingen "evolution" som leder till uppkomsten av nya egenskaper någonsin äger rum där.148
Antibiotikaresistens och DDT immunitet är inte bevis för evolution.
Ett av de biologiska koncept som evolutionister försöker att presentera som bevis på deras teori är resistensen hos bakterier mot antibiotika. Många evolutionistiska källor visar antibiotikaresistens som "ett exempel på utvecklingen av levande ting genom fördelaktiga mutationer". En liknande tillämpning görs också för de insekter som bygger upp en immunitet mot insektsgift som DDT.
Emellertid har evolutionister även här fel.
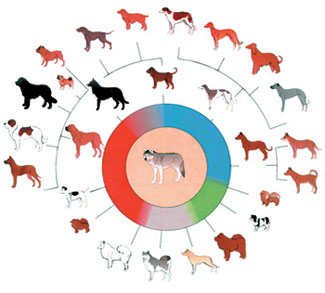
Evolutionister skildrar bakteriernas resistens mot antibiotika som ett bevis för evolution men på ett bedrägligt vis.
Antibiotika är "dödande molekyler" som produceras av mikroorganismer för att bekämpa andra mikroorganismer. Den första antibiotikan var penicillin som upptäcktes av Alexander Fleming 1928. Fleming förstod att mögel som producerade fram en molekyl som dödade Staphylococcus bakterien, och denna upptäckt markerade en vändpunkt i den medicinska världen. Antibiotika som härrör från mikroorganismer användes mot bakterier och resultaten var framgångsrika.
Snart upptäcktes något nytt. Över en tidsperiod bygger bakterier upp immunitet mot antibiotika. Mekanismen fungerar så här: En stor del av de bakterier som utsätts för antibiotika dör, men vissa andra, som inte berörs av detta antibiotikum, replikeras snabbt och utgör snart hela bakteriepopulationen. Således blir hela populationen immun mot antibiotika.
Evolutionister försöker att presentera detta som "bakteriers evolution genom anpassning till villkor".
Sanningen är dock mycket annorlunda än den här ytliga tolkningen. En av de forskare som har gjort den mest detaljerade forskningen inom detta område är den israeliska biofysikern Lee Spetner, som även är känd för sin bok Not by Chance som publicerades 1997. Spetner vidhåller att immuniteten hos bakterier kommer till stånd genom två olika mekanismer, men ingen av dem utgör ett bevis för evolutionsteorin. Dessa två mekanismer är:
1) Överföringen av resistensgener som redan finns bevarade i bakterier.
2) Uppbyggnaden av resistens som en följd av att förlora genetisk information på grund av mutation.
Professor Spetner förklarar den första mekanismen i en artikel publicerad år 2001:
Vissa mikroorganismer är utrustade med gener som medger resistens mot dessa antibiotika.
Resistensen kan vara antingen i form av nedbrytning av antibiotika molekylen eller att avstötning av den från cellen ... De organismer som har dessa gener kan överföra dem till andra bakterier och göra dem resistenta också. Även om resistensmekanismerna är specifika för ett visst antibiotikum, har de flesta patogena bakterier ... lyckats ackumulera ett flertal uppsättningar av gener vilket ger dem resistens mot varierande antibiotika.149
Spetner fortsätter sedan med att säga att detta inte är "bevis för evolution":
Förvärvet av antibiotikaresistens på detta sätt ... är inte av den typ som kan fungera som en prototyp för de mutationer som behövs för evolution. De genetiska förändringar som kan illustrera teorin ska inte enbart lägga till information till bakteriens arvsmassa, de måste lägga till ny information till biokosmos. Den horisontella överföringen av gener sprider bara runt gener som redan finns i vissa arter.150
Vi kan alltså inte tala någon evolution här, eftersom ingen ny genetisk information produceras: genetisk information som redan finns överförs helt enkelt mellan bakterier.
Den andra formen av immunitet, som uppkommer som ett resultat av mutation, är inte heller ett exempel på evolution. Spetner skriver:
... En mikroorganism kan ibland utveckla resistens mot ett antibiotikum genom en slumpmässig substitution av en enda nukleotid ... Streptomycin, som upptäcktes av Selman Waksman och Albert Schatz och refererades första gången 1944, är ett antibiotikum mot vilket bakterier kan utveckla motstånd på detta sätt. Men trots att mutationen som de genomgår i processen är fördelaktig för mikroorganismen i närvaro av streptomycin, kan den inte tjäna som prototyp för den typ av mutationer som erfordras av NDT [Neo Darwinian Theory]. Den typ av mutation som ger resistens mot streptomycin visar sig i ribosomen och försämrar dess molekylära överensstämmelse med antibiotika molekylen. Denna förändring i ytan av mikroorganismens ribosom förhindrar streptomycin molekylen från att fästa och utföra sin antibiotiska funktion. Det visar sig att denna nedbrytning är en förlust av specificitet och därmed en förlust av information. Den viktigaste punkten är att evolution inte kan uppnås genom mutationer av detta slag, oavsett hur många av dem det finns. Evolution kan inte byggas genom att ackumulera mutationer som endast försämrar specificiteten.151
För att sammanfatta: En mutation som gör intrång på en bakteries ribosomer gör bakterien resistent mot streptomycin. Anledningen till detta är "nedbrytningen" av ribosomen genom mutation. Det vill säga, ingen ny genetisk information tillförs bakterien. Tvärtom är ribosomens struktur sönderdelad, det vill säga, bakterien mister sin förmåga att fungera.
(Dessutom har det upptäckts att den muterade bakteriens ribosom är mindre funktionell än den hos en normal bakterie). Eftersom detta "funktionshinder" förhindrar antibiotikan från att fästa på ribosomen utvecklas "antibiotikaresistens".
Slutligen finns det inget exempel på mutation som "utvecklar den genetiska informationen".
Samma sak som ovan nämnts gäller för den immunitet som insekter utvecklar mot DDT och liknande insektsmedel. I de flesta av dessa fall används immunitets gener som redan finns. Den evolutionistiska biologen Francisco Ayala medger detta faktum genom att säga: "De genetiska varianter som krävs för resistens mot de mest skilda typer av bekämpningsmedel var uppenbarligen närvarande i var och en populationen som utsattes för dessa konstgjorda ämnen”.152
Några andra exempel som förklaras av mutation, precis som med den tidigare nämnda ribosommutationen, är fenomen som orsakar "genetisk informations underskott" hos insekter.
I detta fall kan man inte hävda att immunitetsmekanismerna i bakterier och insekter utgör bevis för evolutionsteorin. Det beror på att evolutionsteorin bygger på påståendet att levande varelser utvecklas genom mutationer. Spetner förklarar att varken immunitet mot antibiotika eller några andra biologiska fenomen indikerar ett sådant exempel på mutation, han säger: ”De mutationer som behövs för makroevolution har aldrig observerats, inga slumpmässiga mutationer som kunde representera de mutationer som krävs i NDT som har undersökts på molekylär nivå har tillfört någon information”. Den fråga jag ställer är: Är de mutationer som har observerats av den typ som teorin behöver såsom stöd? Svaret visar sig vara NEJ!”.154
Felaktigheten med rudimentära organ.
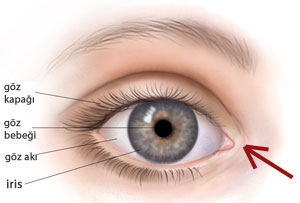
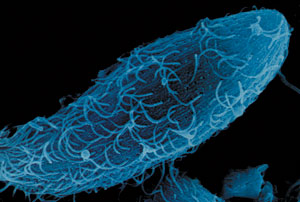
Under tiden som har gått har alla hävda förekomster av rudimentära organ motbevisats. Till exempel det halvmånformade vecket i ögat, omnämnd i The Origins som ett rudimentärt organ. Den har i vår tid visat sig vara fullt fungerande, även om dess funktion var okänd under Darwins tid. Detta organ smörjer ögongloben.
Under lång tid förekom begreppet "rudimentära organ" ofta i den evolutionistiska litteraturen som "bevis" för evolution. Så småningom lades det, i tysthet, åt sidan när det visade sig vara ogiltigt. Men vissa evolutionister tror fortfarande på det, och från tid till annan kommer någon fram som söker framlägga "rudimentära organ" som viktiga bevis för evolution.
Begreppet "rudimentära organ" lades fram första gången för ett sekel sedan. Evolutionister gjorde gällande att det i vissa varelsers kroppar fanns ett antal icke-funktionella organ. De hade gått i arv från deras stamfäder och hade så småningom blivit rudimentära av brist på nyttjande.
Hela antagandet är ganska ovetenskapligt, och bygger helt på otillräcklig kunskap. Dessa "icke-funktionella organ" var i själva verket organ vars "funktion ännu inte hade upptäckts". Den bästa indikationen på detta var den gradvisa men betydande minskningen av rudimentära organ på evolutionisternas långa lista. S. R. Scadding, själv en evolutionist, instämde med detta faktum i sin artikel "Can vestigial organs constitute evidence for evolution?” som publicerades i tidskriften Evolutionary Theory:
Eftersom det inte är möjligt att entydigt identifiera onödiga strukturer och eftersom den struktur som används i argumentet inte är vetenskapligt giltig, drar jag slutsatsen att "rudimentära organ" inte ger något särskilt bevis för teorin om evolution.155
Listan över rudimentära organ som sammanställdes av den tyske anatomen R. Wiedersheim år 1895 omfattade cirka 100 organ, inklusive blindtarmen och svanskotan. Allteftersom vetenskapen fortskred upptäcktes det att alla organ i Wiedersheims lista faktiskt hade mycket viktiga funktioner. Till exempel upptäcktes det att blindtarmen, som skulle vara ett "rudimentärt organ", i själva verket var ett lymfatiskt organ som bekämpade infektioner i kroppen. Detta faktum klargjordes 1997: "Andra organ och vävnader i kroppen, thymus (brässen), levern, mjälten, blindtarmen, benmärgen, och de små ansamlingarna av lymfatisk vävnad såsom tonsillerna i halsen och Peyers plaque i tunntarmen utgör också en del av det lymfatiska systemet, De hjälper också kroppen att bekämpa infektioner”156
Det upptäcktes även att tonsillerna, som ingick i samma lista över rudimentära organ, hade en betydande roll för att skydda halsen mot infektioner, i synnerhet hos barn fram till pubertetsåldern. Man fann att svanskotan vid den nedre änden av ryggraden stödjer skelettet kring bäckenet och är konvergenspunkt för vissa små muskler och av denna anledning skulle det inte vara möjligt att sitta bekvämt utan svanskotan. Under åren som följde, insåg man att brässen utlöste immunsystemet i människokroppen genom att aktivera de så kallade T-cellerna, att tallkottkörteln var ansvarig för utsöndringen av några viktiga hormoner, att sköldkörteln var effektivt för att ge jämn tillväxt hos spädbarn och barn, och att hypofysen kontrollerar funktionen hos många hormonella körtlar. Alla dessa var en gång betraktade som "rudimentära organ". Slutligen har det halvmånformade vecket i ögat, som kallades ett rudimentär organ av Darwin, i själva verket påvisats ha ansvar för rengöring och smörjning av ögongloben.
Det fanns ett mycket viktigt logiskt fel i evolutionistiska hävdandet om rudimentära organ. Som vi har konstaterat var detta hävdande att rudimentära organ hos levande ting gick arv från deras förfäder.
Däremot har vissa av de påstådda "rudimentära" organen inte funnits i de arter som påstås vara människans förfäder! Till exempel existerar inte blindtarmen hos vissa arter av apor som sägs vara människans förfäder. Den berömda biologen H. Enoch, som utmanade teorin om rudimentära organ, uttryckte detta logiska fel som följer:
Apor har en blindtarm, medan deras mindre nära släktingar, halvaporna, inte har det; men det verkar återigen finnas bland de ännu lägre däggdjuren såsom opossum. Hur kan evolutionisterna förklara detta?157
Enkelt uttryckt innehåller scenariot om rudimentära organ som har lagts fram av evolutionister en rad allvarliga logiska brister, och har i varje fall visat sig falskt ur vetenskaplig synvinkel. Det finns inte ett enda ärftligt rudimentärt organ i människokroppen, eftersom människan inte utvecklades från andra varelser som en följd av slumpen, men skapades i sin nuvarande, kompletta och perfekta form.
Myten om homologi.
Inom biologin kallas strukturella likheter mellan olika arter "homologi". Evolutionister försöker framlägga dessa likheter som bevis för evolution.
Darwin trodde att varelser med liknande (homologa) organ hade ett evolutionärt förhållande till varandra, och att dessa organ måste ha ärvts från en gemensam förfader. Enligt hans antagande, hade både duvor och örnar vingar, och därför hade duvor, örnar, och faktiskt alla andra fåglar med vingar förment utvecklats från en gemensam förfader.



Kartal, yarasa ve sinek, üçü de kanatlara sahip canlılardır. Ancak onların benzer organlara sahip olmaları, ortak bir atadan evrimleştiklerinin delili değildir.
Homologi är ett bedrägligt argument framförd på grundval av inga andra bevis än en uppenbar fysisk likhet. Detta argument har aldrig verifierats av en enda konkret upptäckt under alla år sedan Darwins tid. Ingenstans i världen har någon kommit fram med ett fossil av den inbillade gemensamma förfadern till varelser med homologa strukturer. Dessutom klargör följande punkter att homologi inte ger några belägg för att evolution någonsin inträffat.
1. Man finner homologa organ i varelser som tillhör helt olika phyla, bland vilka evolutionister inte har kunnat fastställa någon form av evolutionärt släktskap;
2. De genetiska koderna hos vissa varelser som har homologa organ är helt olika varandra.
3. Den embryologiska utvecklingen av homologa organ i olika varelser är helt annorlunda.
Låt oss nu undersöka var och en av dessa punkter en efter en.
Liknande organ hos helt olika levande arter.
Däggdjurs tvillingar som trotsar homologi
TVÅ OBESLÄKTADE, UTDÖDA DÄGGDJUR MED JÄTTELIKA TÄNDER
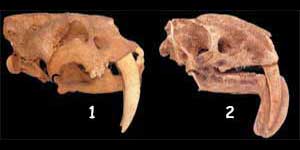
Ett annat exempel på extraordinär likhet mellan däggdjur med moderkaka (placenta) respektive pungdäggdjurs ”tvillingar” är den mellan de utdöda däggdjuren Smilodon (nedan) och Thylacosmilus (ovan), som bägge var rovdjur med enorma framtänder. Den stora graden av likhet mellan skallen och tandstruktur hos dessa två däggdjur, mellan vilka inget evolutionärt släktskap kan fastställas, välter den homologiska uppfattningen att liknande strukturer är bevis till förmån för evolution.
TASMANSKA VARGEN OCH DESS NORDAMERIKANSKA MOTPART


Närvaron av tvilling arter mellan pungdjur och placenta däggdjur utdelar ett dråpslag mot hävdandet om homologi. Till exempel: Den tasmanska vargen (ett pungdjur) och vargen med moderkaka som återfinns i Nordamerika liknar varandra till en anmärkningsvärd nivå. Ovan syns skallarna av dessa två mycket snarlika djur. En sådan likhet mellan de två, som inte kan antas ha något evolutionärt släktskap ogiltigförklarar homologihävdandet
Det finns ett antal homologa organ i olika grupper bland vilka evolutionister inte kan fastställa någon form av evolutionärt släktskap. Vingarna är ett exempel. Förutom fåglar, finner vi vingar på fladdermöss, som är däggdjur och insekter och även på vissa dinosaurier som är utdöda reptiler. Inte ens evolutionister sätter ett evolutionärt släktskap eller familjerelation mellan dessa fyra olika grupper.
Ett annat slående exempel är den fantastiska likheten och den strukturella likheten som observeras i ögonen på olika varelser. Till exempel är bläckfisken och människan två mycket olika arter, mellan vilka ingen evolutionär relation någonsin torde föreslås, men ögonen hos bägge är mycket lika varandra i fråga om struktur och funktion. Inte ens evolutionister försöker påtala likheten mellan ögonen hos bläckfisken och människan genom att sätta dem under en gemensam förfader. Dessa och många andra exempel visar att evolutionisternas hävdande som grundar sig på likheter är helt ovetenskapligt.
I själva verket borde homologa organ vara en mycket pinsamma för evolutionister. Den berömda evolutionisten Frank Salisburys bekännelser avslöjade hur extremt olika varelser kom att få mycket likartade ögon understryker en återvändsgränd av homologi:
Även något så komplext som ögat har dykt upp flera gånger, till exempel hos bläckfisken, hos ryggradsdjuren och hos leddjuren. Det är illa nog att stå för ursprunget av sådana saker en gång, men tanken att framhäva dem flera gånger enligt den moderna syntetiska teorin får tanken att svindla.158
Det finns många varelser som, trots en mycket likartad fysisk uppbyggnad, inte tillåter några påståenden om evolutionär släktskap. Två stora däggdjurs kategorier placentaler och pungdjur, är ett exempel. Evolutionister anser att denna åtskillnad uppstod när däggdjur först dök upp, och att varje grupp levde sin egen evolutionära historia helt oberoende av den andra. Men det är intressant att det finns "par" i placentaler och pungdjur som är nästan likadana. De amerikanska biologerna Dean Kenyon och Percival Davis gör följande kommentar: Enligt Darwins teori utvecklades mönstret för vargar, katter, ekorrar, svin, myrslokar,mullvadar, och möss två gånger:
en gång i moderkakan hos däggdjur och återigen, helt oberoende, i pungdjur. Detta innebär att det häpnadsväckande påståendet om slumpmässig oriktad mutation och urvals processer pånågot sätt träffade identiska karaktäristika flera gånger i vitt skilda organismer.159


När det gäller struktur, är ögonen hos människor och bläckfiskar mycket lika varandra. Men det faktum att de två arterna har liknande organ innebär inte att de har utvecklats från en gemensam förfader. Inte ens evolutionister försöker hänvisa till likheten i ögonens struktur hos bläckfisken och människan genom att anslå en gemensam förfader.
.
Extraordinära likheter och liknande organ som dessa, som evolutionistiska biologer inte kan acceptera som exempel på "homologi", visar att det inte finns något bevis för tesen om utvecklingen från en gemensam förfader. Vad kan då vara en vetenskaplig förklaring till dessa liknande strukturer hos levande varelser? Svaret på den frågan gavs innan Darwins evolutionsteori kom att dominera den vetenskapliga världen. Vetenskapsmän som Carl von Linné, som först systematiserade levande organismer i enlighet med deras liknande strukturer, och Richard Owen betraktade dessa strukturer som exempel på en vanlig form av skapelse. Med andra ord liknande organ (eller, numera, liknande gener) anses finnas eftersom de skapades för att tjäna ett visst syfte, inte för att de hade utvecklats av en slump från en gemensam förfader.
Moderna vetenskapliga rön visar att påståendet om en "gemensam förfader" ställd med hänsyn till liknande organ är felaktig, och att den enda möjliga förklaringen är vanlig skapelse, bekräftar ännu en gång att levande varelser skapades av Allah.
Den genetiska och embryologiska återvändsgränden av homologi.
För att evolutionisternas påstående om att "homologi" skall tas på allvar, bör liknande (homologa) organ hos olika varelser också kodas med liknande (homologa) DNA-koder. Så är det emellertid inte. Liknande organ styrs ofta av mycket olika genetiska (DNA) koder. Dessutom är liknande genetiska koder i DNA hos olika djur ofta förknippade med helt olika organ.

Professor Michael Denton: "Evolution är en teori i kris"
Michael Denton, en professor i biokemi från Australien, beskriver i sin bok Evolution: A Theory in Crisis den genetiska återvändsgränden av evolutionisternas tolkning av homologi:
"Homologa strukturer är ofta specificerade av icke-homologa genetiska system och begreppet homologi kan sällan återföras till embryologi”.160
Ett berömt exempel på detta ämne är den "femfingriga skelettstrukturen" hos fyrfotadjur, som citeras i nästan alla evolutionistiska läroböcker. Fyrfotadjur, det vill säga landlevande ryggradsdjur, har fem fingrar/tår på sina fram respektive bakben.Även om dessa inte alltid har utseendet av fem fingrar/tår såsom vi känner igen dem, räknas de är alla som pentadactyla (femfingriga) på grund av benstommen. Fram och bakbenen av en groda, en ödla, en ekorre eller en apa har alla samma struktur. Även benstrukturen hos fåglar och fladdermöss överensstämmer med denna grundläggande struktur.
Evolutionister hävdar att alla levande varelser härstammar från en gemensam förfader, och de har länge hänvisat till lemmar av pentadactyl form som bevis på detta. Detta påstående nämndes i nästan alla grundläggande källor i biologi under hela 1900-talet som mycket starka bevis för evolution. Genetiska fynd på 1980-talet motsäger detta evolutionistiska påstående. Man insåg att ett pentadactyl lemmönster hos olika varelser styrs av helt olika gener. Den evolutionistiska biologen William Fix beskriver kollapsen av evolutionisternas tes om pentadactylism på detta sätt:
De äldre läroböckerna om evolution gör mycket av idén kring homologi genom att påpeka de uppenbara likheterna mellan skelettet i armar och ben hos olika djur. Således återfinns "pentadactyl" lemmönstret i människans arm, i fågelns vinge, och bröstfenan hos en val, och detta anses indikera deras gemensamma ursprung. Om nu dessa olika strukturer hade överförts av samma gen par, varierade från tid till annan genom mutationer och detta hade skett genom miljöurvalet skulle teorin vara meningsfull. Tyvärr är detta inte fallet. Homologa organ uppstår urhelt olika genkomplex i de olika arterna. Konceptet om att homologi i termer av liknande gener som går i arv från en gemensam förfader har fallit sönder ... 161
En annan sak är den att för att den evolutionära avhandlingen om homologi skall tas på allvar, behöver perioderna för liknande strukturers embryologiska utveckling, det vill säga: de olika stadierna av utveckling i ägget eller moderns livmoder, behöva vara parallella men i verkligheten är dessa embryologiska perioder för liknande strukturer helt olika från art till art.
Avslutningsvis kan vi säga att genetisk och embryologisk forskning har visat att begreppet homologi som definieras av Darwin som "bevis på utvecklingen av levande varelser från en gemensam förfader" inte kan utgöra något bevis alls. I detta avseende kan vetenskapen sägas ha bevisat gång på gång att den darwinistiska tesen är falsk.
Ogiltigheten av hävdandet om molekylär homologi.
Evolutionisters befordran av homologi som bevis för evolution är ogiltigt inte enbart på morfologisk nivå utan även på molekylär nivå. Evolutionister säger att DNA-koderna, eller motsvarande protein strukturer, hos olika levande arter är likartade, och att det i denna likhet finns belägg för att dessa levande arter har utvecklats från gemensamma förfäder eller från varandra. I verkligheten fungerar dock inte resultaten av molekylära jämförelser till förmån för evolutionsteorin alls. Det finns stora molekylära skillnader mellan varelser som verkar vara väldigt lika och relaterade. Till exempel är cytokrom-C proteinet, ett av de proteiner som är viktiga för andningen, mycket olika hos levande varelser av samma klass. Enligt forskning som bedrivs i kring detta är skillnaden mellan två olika kräldjurs arter större än skillnaden mellan en fågel och en fisk eller en fisk och ett däggdjur. En annan studie har visat att molekylära skillnader mellan vissa fågelarter är större än skillnaderna mellan samma fågelarter och däggdjuren. Det har även upptäckts att den molekylära skillnaden mellan bakterier som verkar vara mycket lika är större än skillnaden mellan däggdjur, amfibier och insekter.162Liknande jämförelser har gjorts med hemoglobin, myoglobin, hormoner, och gener och man har kommit fram till liknande slutsatser.163Beträffande dessa fynd inom molekylär biologi, kommentarer Dr Michael Denton:
Varje klass på en molekylär nivå är unik, isolerad och olänkad av intermediärer. Molekyler har således, liksom fossiler, misslyckats att tillhandahålla de svårfångade mellanprodukter som så länge har efterfrågats av evolutionär biologi ... På molekylär nivå är ingen organism "förfader" eller "primitiv" eller "avancerad" jämfört med sina släktingar ... Det råder inget tvivel om att om detta molekylära bevis hade funnits för hundra år sedan hade tanken på organisk evolution kanske aldrig blivit accepterad.164
Kollapsen av "Livets träd".
Under 1990-talet förvärrade forskningen om levande tings genetiska koder det dilemma som evolutionsteorin står inför i detta avseende. I dessa experiment jämfördes sekvenser av "ribosomalt RNA" (rRNA), i stället för de tidigare jämförelserna som var begränsade till proteinsekvenser. Från dessa resultat, försökte evolutionistiska forskare att etablera ett "evolutionärt träd".
Emellertid blev de besvikna över resultaten. Enligt en artikel år 1999 av de franska biologerna Hervé Philippe och Patrick Forterre, ”Med fler och fler sekvenser tillgängliga, visade det sig att de flesta proteiners fylogenier motsäger varandra liksom rRNA trädet”.165
Förutom rRNA jämförelser jämfördes även DNA-koderna i generna hos levande varelser, men resultaten har varit motsatsen till "livets träd" som förutsätts av evolution.
Molekylärbiologerna James A. Lake, Ravi Jain och Maria C. Rivera nystar vidare på detta i en artikel år 1999:
Forskare började analysera en mängd gener från olika organismer och fann att deras förhållande till varandra motsade att det evolutionära livets träd härrör enbart från rRNA analysen.166
Varken jämförelser som har gjorts av proteiner, rRNA eller gener, bekräftar premisserna av evolutionsteorin. Carl Woese, en välrenommerad biolog från University of Illinois medger att begreppet "fylogeni" har förlorat sin betydelse mot bakgrunden av molekylära fynd på detta sätt:
”Ingen konsekvent fylogeni har vuxit fram ur många enskilda proteinernas fylogenier som hittills har framställts och fylogenetiska missförhållanden kan ses överallt i det universella trädet, från dess rot till de stora förgreningarna inom och mellan de olika grupperna till strukturen av de enskilda primära grupperingarna”.167
Det faktum att resultatet av molekylära jämförelser inte talar för, utan snarare emot, evolutionsteorin medgavs också i artikeln som heter "Is it time to uproot the Tree of Life?”, och som publicerades i Science år 1999. I denna artikel av Elizabeth Pennisi uppges att de genetiska analyser och jämförelser som utfördes av darwinistiska biologer för att belysa "livets träd" faktiskt gav direkt motsatta resultat och hon fortsätter med att säga att "nya uppgifter grumlar den evolutionära bilden":
För ett år sedan, när biologer granskade nyligen sekvenserade genom från mer än ett dussin mikroorganismer trodde de att dessa data kunde stödja den accepterade kartläggningen av livets tidiga historia. Men vad de såg förvirrade dem. Jämförelser av de då tillgängliga genomen underlät inte enbart att förtydligade bilden av hur livets stora grupperingar utvecklats, de förvirrade den. Och nu, med ytterligare åtta mikrobiella sekvenser till hands, har situationen blivit ännu mer förvirrande.. Många evolutionära biologer hade antagit att de, på ett grovt sätt, kunde urskilja början till livets tre riken av levande ting (som är djurriket, växtriket och riket för encelliga livsformer) ... När hela DNA-sekvenser hade öppnat vägen för att jämföra andra typer av gener, förväntade sig forskare att de helt enkelt skulle utöka trädets detaljrikedom. Men "ingenting kunde ha varit mer felaktigt", säger Claire Fraser, chef för Institutet för genomforskning (TIGR) i Rockville, Maryland. I stället har jämförelserna gett många versioner av livets träd som skiljer sig från rRNA trädet och även är i inbördes konflikt med varandra... 168
Kort sagt, allteftersom molekylär biologiskt fortskrider, förlorar homologikonceptet mer mark. Jämförelser som gjorts av proteiner, rRNA och gener visar att varelser som påstås vara nära släktingar enligt evolutionsteorin faktiskt är helt skilda från varandra. En studie från 1996 med 88 proteinsekvenser grupperade kaniner med primater i stället för gnagare, en analys 1998 av 13 gener i 19 djurarter placerade sjöborrar bland Chordata, och en annan studie 1998 baserad på 12 proteiner satte kor närmare valar än hästar.Molekylärbiolog Jonathan Wells sammanfattade år 2000 situationen på följande sätt: Inkonsekvenser bland släktträd baserade på olika molekyler, och de bisarra släktträd som resulterar från vissa molekylära analyser, har nu försänkt molekylär fylogeni i en kris 169
"Molekylär fylogeni" står inför en kris, vilket innebär att evolutionsteorin också står inför en kris. Fylogeni hänvisar till så kallade familjerelationer "mellan olika levande ting och är den hypotetiska grunden för teorin om evolutionen”. Återigen, undergräver vetenskapen tesen att levande varelser utvecklats från varandra, vilket visar att alla levande grupper skapades separat.
Myten om embryologisk rekapitulering.

Haeckel var en evolutionist som var ännu mer nitisk än Darwin i många avseenden. Av den anledningen tvekade han inte att förvränga vetenskapliga fakta och ta fram olika förfalskningar.
Vad som förr kallas "rekapituleringsteorin" har länge varit eliminerad från vetenskaplig litteratur, men är fortfarande presenterat som en vetenskaplig realitet i några evolutionistiska publikationer. Termen "rekapitulation" är en kondensering av satsen "ontogeni rekapitulerar fylogeni", som lades fram av den evolutionistiske biologen Ernst Haeckel i slutet av 1800-talet.
Denna teori av Haeckel förutsätter att levande embryon återupplever den evolutionära process som deras pseudoförfäder genomgick. Han teoretiserade att under sin utveckling i livmodern, uppvisar det mänskliga embryot först egenskaper hos en fisk, och sedan de av en reptil, och slutligen de av en människa.
Det har sedan visat sig att denna teori är helt falskt. Det är nu känt att "gälarna" som visar sig i ett tidigt skede av det mänskliga embryot i själva verket är de inledande faserna av mellanörat, bisköldkörteln och thymus. Den del av embryot som liknar "gulesäcken" visar sig vara en säck som producerar blod för spädbarnet. Den del som hade identifierats som en "svans" av Haeckel och hans anhängare är i själva verket ryggraden, som liknar en svans bara för att det tar form innan benen gör.
Dessa utgör universellt erkända fakta i den vetenskapliga världen och accepteras även av evolutionisterna själva. George Gaylord Simpson, en av grundarna av neo-darwinism, skriver:
Haeckels tog miste på den involverade evolutionära principen. Det är nu fastställt att ontogeni inte upprepar fylogeni.171I en artikel publicerad i American Scientist, läser vi:Säkerligen är den biogenetiska lagen stendöd. Det uteslöts slutligen ur läroböcker i biologi på femtiotalet. Som ett ämne för allvarliga teoretiska utredningar var den utrotad under tjugotalet ... 172
En annan intressant aspekt av "rekapitulation" var Ernst Haeckel själv, en förfalskare som förfalskade sina teckningar för att stödja teorin som han förespråkade. Haeckels förfalskade påståenden för att visa att fiskar och mänskliga embryon liknade varandra. När han avslöjades, var det enda försvar han erbjöd den att andra evolutionister hade begått liknande brott:
HAECKELS BEDRÄGLIGA TECKNINGAR
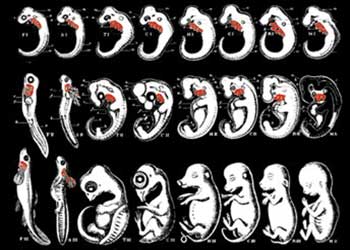
Dessa teckningar fabricerades av Haeckel för att framhäva "likheterna" mellan mänskliga och fisk embryon. Jämförande av hans skiss med ett äkta mänskligt embryo visar att han medvetet har utelämnat en stor del av de faktiska organen. (Francis Hitching, The Neck of the Giraffe. Where Darwin Went Wrong sid 205)
Efter denna blottställande bekännelse av ”urkundsförfalskning" borde jag anse mig fördömd och förintad om jag inte hade trösten att vid min sida på den anklagades bänk se hundratals andra syndare, bland dem många av de mest betrodda observatörer och mest uppskattade biologer. Den stora majoriteten av alla diagram i de bästa biologiska läroböcker, avhandlingar och tidskrifter är i samma grad ansvariga för "urkundsförfalskning", för alla är inexakta, och är mer eller mindre tillrättalagda, schematiserade och konstruerade.173
Det finns verkligen "hundratals andra syndare, bland dem många av de mest betrodda observatörerna och mest uppskattade biologerna" vars studier är fulla av fördomsfulla slutsatser, förvrängningar, och även förfalskningar. Detta eftersom de alla har betingat sig att kämpa för evolutionsteorin även om det inte finns tillstymmelse till vetenskapliga bevis som stöder den.
DİPNOTLAR
143. Loren Eiseley, The Immense Journey, Vintage Books, 1958, sid 186
144. Charles Darwin, The Origin of Species: A Facsimile of the First Edition, Harvard University Press, 1964, sid 184
145. Norman Macbeth, Darwin Retried: An Appeal to Reason, Harvard Common Press, New York: 1971, sid 33
146. Norman Macbeth, Darwin Retried: An Appeal to Reason, sid 36
147. Loren Eiseley, The Immense Journey, Vintage Books, 1958. s 227
148. Ayrıntılı bilgi için bkz. Jonathan Wells, Icons of Evolution, 2000, sid 159-175
149. Dr. Lee Spetner, "Lee Spetner/Edward Max Dialogue: Continuing an exchange with Dr. Edward E. Max", 2001, http://www.trueorigin.org/spetner2.asp
150. Dr. Lee Spetner, http://www.trueorigin.org/spetner2.asp
151. Dr. Lee Spetner, http://www.trueorigin.org/spetner2.asp
152. Dr. Lee Spetner, http://www.trueorigin.org/spetner2.asp
153. Francisco J. Ayala, "The Mechanisms of Evolution", Scientific American, cilt 239, Eylül 1978, sid 64
154. Dr. Lee Spetner, http://www.trueorigin.org/spetner2.asp
155. sid R. Scadding, "Do 'Vestigial Organs' Provide Evidence for Evolution?", Evolutionary Theory, Cilt 5, Mayıs 1981, sid 173
156. The Merck Manual of Medical Information, Home edition, New Jersey: Merck & Co., Inc. The Merck Publishing Group, Rahway, 1997
157. H. Enoch, Creation and Evolution, New York: 1966, sid 18-19
158. Frank Salisbury, "Doubts About the Modern Synthetic Theory of Evolution", American Biology Teacher, Eylül 1971, sid 338
159. Dean Kenyon & Percival Davis, Of Pandas and People: The Central Question of Biological Origins (Dallas: Haughton Publishing, 1993), sid 33
160. Michael Denton, Evolution: A Theory in Crisis, London, Burnett Books, 1985, sid 145
161. Fix, William, The Bone Peddlers: Selling Evolution (New York: Macmillan Publishing Co., 1984), sid 189
162. W. R. Bird, The Origin of Species Revisited, Thomas Nelson Co., Nashville: 1991, ssid 98-99; Percival Davis, Dean Kenyon, Of Pandas and People, Haughton Publishing Co., 1990, sid 35-38
163. W. R. Bird, The Origin of Species Revisited, sid 98-99, 199-202
164. Michael Denton, Evolution: A Theory in Crisis, London: Burnett Books, 1985, sid 290-91
165. Hervé Philippe and Patrick Forterre, "The Rooting of the Universal Tree of Life is Not Reliable", Journal of Molecular Evolution, vol 49, 1999, sid 510
166. James Lake, Ravi Jain ve Maria Rivera, "Mix and Match in the Tree of Life", Science, vol. 283, 1999, sid 2027
167. Carl Woese, "The Universel Ancestor", Proceedings of the National Academy of Sciences, USA, 95, (1998) sid 6854
168. Elizabeth Pennisi, "MICROBES, IMMUNITY, AND DISEASE: Is It Time to Uproot the Tree of Life?" Science, Volume 284, Number 5418, Issue of 21 May 1999, psid 1305-1307
169. Jonathan Wells, Icons of Evolution, Regnery Publishing, 2000, sid 51
170. Jonathan Wells, Icons of Evolution, Regnery Publishing, 2000, sid 51
171. G. G. Simpson, W. Beck, An Introduction to Biology, New York, Harcourt Brace and World, 1965, sid 241
172. Keith sid Thompson, "Ontogeny and Phylogeny Recapitulated", American Scientist, Cilt 76, Mayıs / Haziran1988, sid 273
173. Francis Hitching, The Neck of the Giraffe: Where Darwin Went Wrong, New York: Ticknor and Fields 1982, sid 204