W poprzednich rozdziałach przedstawiliśmy dowody z dziedziny biologii molekularnej i materiały kopalniane, które świadczyły przeciwko teorii ewolucji. Teraz chcielibyśmy zająć się pojęciami z dziedziny biologii, wykorzystywanymi przez ewolucjonistów w celu udowodnienia słuszności ich twierdzeń.
W genetyce określenia „wariacje” używa się dla opisania zdarzenia, w efekcie którego jednostki lub grupy wewnątrz jakiegoś gatunku przyjmują różne cechy charakterystyczne. Wszyscy ludzie na Ziemi są obdarzeni podobnymi informacjami genetycznymi, ale, dzięki wielkości potencjału wariacyjnego, część z nas ma rude włosy lub skośne oczy, inni długie nosy, a jeszcze inni są niskiego wzrostu.
Te wariacje są przez ewolucjonistów używane w celu udowodnienia słuszności ich teorii. Nie stanowią one jednak żadnego potwierdzenia, ponieważ są tylko wynikiem różnych kombinacji istniejących informacji, a nie zmianą w materiale genetycznym.
Wariacje wewnątrz gatunku nie są dowodem słuszności teorii ewolucji
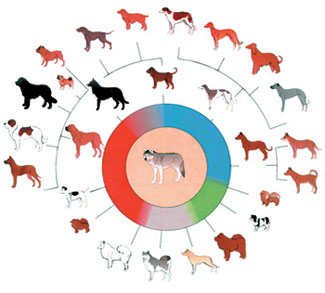
W dziele O powstawaniu gatunków Darwin pomylił dwa różne pojęcia: zmiany wewnątrz jednego gatunku i powstawanie nowego. Obserwując różnice wśród rozmaitych ras psów, twierdził on, że wariacje, które do nich doprowadziły, doprowadzą też do powstania nowego gatunku. Do dziś ewolucjoniści uważają zmiany wewnątrzgatunkowe za dowód słuszności teorii ewolucji i działania doboru naturalnego.
Nie są to jednak dobre argumenty. Nieważne, ile ras psów istnieje na świecie; zawsze pozostaną one psami, nigdy nie staną się innym gatunkiem.
Wariacje odbywają się zawsze w granicach istniejącej informacji genetycznej. W genetyce określa się to mianem „gen-pool”. Wszystkie cechy charakterystyczne danego gatunku mogą ujawniać się na różne sposoby. W wyniku wariacji może pojawić się grupa gadów posiadająca długie ogony i krótkie nogi; zmiany takie jednak nigdy nie dodadzą gadom skrzydeł, nie zmienią ich układu oddechowego czy kostnego i nie przekształcą gada w ptaka. Podobne wariacje wymagałyby dodania nowej informacji genetycznej, a to nie jest w tym przypadku możliwe.
Darwin nie zdawał sobie z powyższego sprawy, kiedy tworzył teorię ewolucji. Myślał on, że wariacje nie mają granic. W jednym z artykułów z 1844 roku pisał: „Fakt, że wariacje w naturze mają pewne granice, przyjęty jest przez większość autorów, ja jednak nie widzę żadnych podstaw, na których opierają oni swoje założenia.”143 W dziele O powstawaniu gatunków Darwin podaje wiele przykładów takich zmian w celu udowodnienia słuszności teorii ewolucji.
Według niego, hodowcy zwierząt, którzy krzyżują ze sobą różne rasy krów w celu podniesienia ich wydajności, uzyskają w końcu zupełnie inny gatunek. Ideę o nieograniczonych wariacjach widać najlepiej w poniższym cytacie z dzieła Darwina:
„Nie widzę problemu w tym, żeby rasa niedźwiedzi, która dzięki selekcji naturalnej przyswoiła sobie cechy zwierząt wodnych, zmieniła się w końcu w stworzenia podobne do wieloryba.”144
Darwin mógł przytoczyć taki przykład, ponieważ nauka w XIX wieku stała jeszcze na bardzo niskim poziomie. Dopiero w zeszłym stuleciu, w wyniku doświadczeń i obserwacji, powstało prawo genetycznej homeostazy. Utworzenie owego prawa było efektem podejmowanych bezskutecznie prób skrzyżowania ze sobą różnych gatunków. Wniosek, do jakiego doszli naukowcy, jest taki, że pomiędzy gatunkami istnieje niemożliwa do przekroczenia bariera. Oznacza to, że hodowcy bydła, krzyżujący różne jego rasy w celu podniesienia wydajności krów, nigdy nie stworzyliby nowego gatunku, jak wydawało się Darwinowi.
Norman Macbeth, który w swojej książce Darwin Retired pokazuje błędy darwinizmu, pisze:
Czy wieloryby pochodzą od niedźwiedzi?

Według Darwina jest to możliwe, gdyż stwierdził on: „Nie widzę problemu w tym, żeby rasa niedźwiedzi, która dzięki selekcji naturalnej przyswoiła sobie cechy zwierząt wodnych, zmieniła się w końcu w stworzenia podobne do wieloryba.”
„Kwintesencję problemu stanowi pytanie, czy wariacje wśród organizmów są nieograniczone... Gatunki wydają się stabilne. Znamy wiele przykładów zdumionych hodowców, którzy, krzyżując różne odmiany roślin czy zwierząt, dochodzili do pewnego nieprzekraczalnego punktu. Po dotarciu do niego – gatunki często wracały do stanu początkowego. Mimo wszystkich wysiłków nie udało się wyhodować czarnego tulipana czy niebieskiej róży.”145
Luther Burbank, jeden z najsłynniejszych hodowców na świecie, komentuje to w następujący sposób: „(...) istnieją pewne granice, wewnątrz których krzyżowanie jest możliwe, a te granice z pewnością podlegają jakimś prawom”.146
Duński uczony, W.L. Johansen, pisze:
„Wariacje, tak podkreślane przez Darwina i Wallace’a, są możliwe tylko do pewnego punktu. Nie można ich uważać za powód stałych (ewolucyjnych) zmian.”147
To samo odnosi się do słynnych zaobserwowanych przez Darwina na Galapagos zięb. Badania przeprowadzone w ostatnich latach pokazały, że zmiany, którym poddawały się te ptaki, nie były, jak to udowadniał Darwin, nieograniczone. Czternaście różnych grup zięb zaobserwowanych przez uczonego to wariacje, które mogły parzyć się ze sobą i tworzyć nowe grupy, ale tylko wewnątrz gatunku. Dowiodły tego badania przebywających długo na Galapagos uczonych, Petera i Rosemary Granit.148
Jedną z tez w biologii, wykorzystywaną w celu udowodnienia słuszności teorii ewolucji, jest teza o odporności bakterii na antybiotyki. Prawie wszystkie ewolucjonistyczne źródła cytują ją jako dowód stopniowego rozwoju organizmów, odbywającego się dzięki pozytywnym mutacjom. W ten sam sposób owady nabrały odporności na środki owadobójcze, takie jak np. DDT.
Jednak i tutaj ewolucjoniści nie mają racji.
Evolutionists portray bacteria’s resistance to antibiotics as evidence of evolution—but in a deceptive way.
Bakterie to „mordercze cząsteczki” produkowane przez niektóre mikroorganizmy w celu zabicia innych mikroorganizmów. Pierwszy antybiotyk, penicylinę, odkrył Alexander Fleming w 1928 roku. Fleming stwierdził, że pleśń wytwarza cząsteczkę, która niszczy bakterie Staphylococus; to odkrycie stanowiło punkt zwrotny w dziejach medycyny. Antybiotyki otrzymywane z mikroorganizmów używane były do walki z wirusami; osiągano bardzo pozytywne wyniki.
Z biegiem czasu dokonano jednak kolejnego odkrycia: niektóre bakterie zaczęły wykazywać odporność na antybiotyki. Mechanizm tego uodpornienia jest następujący: za każdym razem, gdy stosowano owe środki, ginęła większość drobnoustrojów, lecz nie wszystkie, które występowały w zakażonym organizmie. Te, które przetrwały, wykazywały genetyczną odporność na antybiotyki, przekazywaną następnym pokoleniom. W ten sposób zwiększała się ilość niepodatnych na lek stworzeń.
Ewolucjoniści przedstawiają ten fakt jako „ewolucję bakterii w wyniku przystosowywania się do panujących warunków”.
Owa ogólnikowa interpretacja jest jednak daleka od prawdy. Jeden z naukowców zajmujących się tym tematem to izraelski biofizyk, Lee Spetner, znany dzięki opublikowanej w 1997 roku książce Not by Chance. Według Spetnera, istnieją dwa powody zyskania przez bakterie odporności na antybiotyki i żaden z nich nie ma nic wspólnego z ewolucją. Owe mechanizmy to:
Pierwszy mechanizm opisuje Spetner w artykule opublikowanym w 2002 roku:
„Niektóre mikroorganizmy mają geny, które uodparniają się na antybiotyki. Rezystencja może polegać na zdegradowaniu cząstki antybiotyku lub wyrzuceniu jej z komórki... Organizmy posiadające takie geny są w stanie przekazywać je innym bakteriom, przez co one także uodparniają się. Nawet jeżeli mechanizmy immunologiczne skierowane są przeciwko jednemu antybiotykowi, większość patogennych bakcyli ma rozmaite rodzaje genów i uodparnia się na różne bakterie.”149
Prof. Spetner tłumaczy ponadto, dlaczego fakt ten nie jest dowodem na ewolucję owych drobnoustrojów:
„Pozyskanie rezystencji na antybiotyki w ten właśnie sposób... nie mogłoby służyć jako prototyp dla mutacji, jakie są konieczne dla wytłumaczenia ewolucji. Genetyczne zmiany, które byłyby może pomocne w udowadnianiu słuszności tej teorii, musiałyby nie tylko wzbogacić w informacje geny bakterii, ale także dodać je do biokosmosu. Horyzontalny przekaz genów może jedynie rozprzestrzeniać i tak istniejącą informację genetyczną.”150
Nie można tu mówić o jakiejkolwiek ewolucji, ponieważ nie powstaje nowa informacja genetyczna. Pomiędzy bakteriami przekazywana jest bowiem ta, która istniała już wcześniej.
Podobnie i drugi wymieniony przez Spetnera mechanizm nie stanowi dowodu na ewolucyjny rozwój tych drobnoustrojów:
„Czasami mikroorganizmy, dzięki przemieszczeniu się jednego nukleotydu, mogą wykazywać odporność na antybiotyk... Streptomycyna, odkryta przez Selmana Waksmana i Alberta Schatza, o czym po raz pierwszy doniesiono w 1944 roku, jest antybiotykiem, na który bakterie uodparniają się w taki właśnie sposób. Jednak pomimo że mutacje, jakim poddawane są bakcyle podczas działania streptomycyny, mają dodatni wpływ na mikroorganizmy, nie można uznać ich za prototyp tych, o których mówią neodarwiniści. Typ mutacji, który uodparnia bakterie na streptomycynę, pojawia się w rybosomie i niszczy przeciwciała cząsteczki antybiotyku.”151
Spetner porównuje tę sytuację do zepsucia się kluczy. Streptomycyna porównana jest do właśnie klucza, który wchodzi do zamka, przyczepia się do rybosomu bakterii i unieszkodliwia go. Mutacje jednak zmieniają kształt rybosomu, przez co antybiotyk ten nie może się do niego dostać. Spetner kontynuuje:
„Jak widać, deformacja w budowie rybosomu jest niczym innym, jak utratą cech specyficznych, co równa się pozbawieniu informacji. Ewolucja nie może opierać się na tego typu mutacjach, niezależnie od tego, jak dużo ich zaistniałoby.”152
Reasumując: mutacja, do jakiej dochodzi w rybosomie bakterii, sprawia, iż uodparnia się ona na działanie streptomycyny. Powodem jest „rozłożenie” rybosomu przez mutację. Oznacza to, że bakterii nie dodaje się nowej informacji genetycznej, wręcz przeciwnie – struktura rybosomu „rozpada się” i bakterie pozbawione zostają swych cech charakterystycznych, przez co stają się bezużyteczne. W ten sposób powstaje „rezystencja na antybiotyk”.
Jak widać, nie jest to przykład mutacji powielającej informacje genetyczne, a twierdząc przeciwnie, ewolucjoniści są w błędzie.
Analogicznie jest z odpornością na środki owadobójcze, takie jak np. DDT, wykazywaną przez niektóre owady. W większości przypadków mamy do czynienia z działaniem istniejących genów odpornościowych. Przyznaje to ewolucjonistyczny biolog, Francisco Ayala: „Genetyczne wariacje potrzebne do rezystencji przeciwko różnym rodzajom środków owadobójczych występowały wszędzie tam, gdzie stosowane były owe syntetyczne środki.”153 Inne przykłady tłumaczenia rezystencji mutacjami, takimi jak wyżej opisana mutacja rybosomów, są raczej przykładami na „zanik informacji genetycznej u owadów”.
Nie należy więc wysnuwać błędnych wniosków, że odporność na antybiotyki i środki owadobójcze u bakterii oraz owadów jest wynikiem procesu ewolucji. Jeszcze raz zacytujmy słowa prof. Spetnera:
„Mutacje, które byłyby konieczne w procesie makroewolucji, nigdy nie zaistniały. Żadna z nich nie była taką, o jakiej mówią neodarwiniści; po dokładnym przebadaniu na płaszczyźnie cząsteczkowej ani jedna mutacja nie spowodowała poszerzenia informacji genetycznej. W takim razie zapytuję: czy typ obserwowanych mutacji może być tym, jakiego potrzebujemy, by udowodnić słuszność teorii ewolucji? Na to pytanie śmiało można odpowiedzieć: nie.”154

Uznana przez Darwina za organ pozbawiony funkcji fałda w formie półksiężyca po wewnętrznej stronie oka ma za zadanie nawılżać i oczyszczać gałkę oczną.
Przez długi czas w literaturze ewolucjonistycznej często pojawiała się koncepcja rudymentacji (uwsteczniania narządów) jako potwierdzenie procesu ewolucji. Kiedy pogląd ten okazał się nieprawdziwy, wokół tematu zaległa cisza. Do dziś jednak od czasu do czasu niektórzy ewolucjoniści próbują pokazać, że rudymentarne organy są ważnym dowodem na słuszność ich teorii.
Idea owa powstała około sto lat temu. Ewolucjoniści przyjęli, że w ciałach niektórych organizmów istnieją pewne organy, które w ogóle nie funkcjonują i niczemu nie służą. Organy owe miały pochodzić od przodków tych istot i stracić swoje funkcje w wyniku nieużywania ich przez całe pokolenia.
Hipoteza ta jest pozbawiona podstaw naukowych. Mowa tu nie o niefunkcjonujących częściach organizmu, ale o takich, których funkcji jeszcze nie odkryto. Najlepszym przykładem jest długa lista organów, przygotowana przez ewolucjonistycznych badaczy. Jeden z ewolucjonistów, S.R. Scadding, jest jednak innego zdania, co przyznaje w artykule: „Czy zanikające organy mogą być dowodem ewolucji?” (“Can vestigal organsconstitute evidence for evolution?”), który opublikowany został w czasopiśmie Evolutionary Theory:
“Niemożność dokładnego zidentyfikowania funkcji zanikających organów i nienaukowość przedstawianych argumentów doprowadzają mnie do przekonania, że zanikające organy nie są w rzeczywistości dowodem na słuszność teorii ewolucji.”155
Zestawienie zanikających organów, przygotowane przez niemieckiego anatoma, R. Wiedersheima, opublikowane w 1895 roku, zawierało około stu pozycji. W tym m.in. takie części, jak: wyrostek robaczkowy, migdałki czy kość ogonowa. Z czasem okazało się, że znajdujące się na liście organy pełnią ważne życiowe funkcje. Wyrostek robaczkowy okazał się zwalczającą infekcje organizmu częścią układu limfatycznego. Fakt ten opisany jest w źródłach medycznych z 1997 roku: „Takie części ciała i tkanki, jak: timus, wątroba, ślepa kiszka, szpik kostny – są ważnymi częściami układu limfatycznego, który służy do zwalczania infekcji i ochrony organizmu.”156
Okazało się także, iż migdałki pełnią ważną rolę (szczególnie u dzieci dojrzewających) w ochranianiu gardła przed infekcjami. Znajdująca się na liście zanikających organów kość ogonowa ma za zadanie wspomaganie miednicy i jest połączeniem dla wielu mniejszych mięśni. Uznana przez Darwina za organ pozbawiony funkcji fałda w formie półksiężyca po wewnętrznej stronie oka – ma za zadanie nawılżać i oczyszczać gałkę oczną.
Tworząc teorię zanikających organów, ewolucjoniści dopuścili się także poważnego błędu logicznego. Jak wcześniej zaznaczyliśmy, uważali oni, że organy te przekazywane były z pokolenia na pokolenie. Jednak w wyniku badań i obserwacji stwierdzono, że niektóre z owych części nie występują u gatunków uważanych przez ewolucjonistów za przodków człowieka. Małpy, na przykład, nie mają wyrostka robaczkowego. Ów logiczny błąd komentuje znany biolog, H. Enoch:
„Małpy człekokształtne mają wyrostek robaczkowy, ale nie występuje on u innych, spokrewnionych z nimi, niższych gatunków tych zwierząt. Organ ów widzimy poza tym u torbaczy (opossum). Jak wytłumaczą ten fakt ewolucjoniści?”157
Krótko mówiąc, próba udowodnienia słuszności teorii ewolucji za pomocą tezy zanikających organów była błędem naukowym i logicznym. W organizmie ludzkim nie ma żadnego zanikającego, pozbawionego funkcji organu odziedziczonego po przodkach. Człowiek nie powstał bowiem przypadkowo, ale w wyniku aktu stworzenia. Pojawił się na Ziemi w swojej ostatecznej i doskonałej formie.
Występowanie podobieństw w budowie narządów u różnych gatunków określa się mianem homologii. Ewolucjoniści próbują przedstawić te zbieżności jako dowód przemawiający za słusznością ich teorii.
Darwin uważał, że stworzenia mające homologiczne organy są ze sobą spokrewnione. Części te bowiem miały zostać odziedziczone po wspólnym przodku. Według Darwina, ponieważ i gołąb, i orzeł mają skrzydła, należy przyjąć, że miały one wspólnego przodka. Więcej: nie tylko gołębia i orła, ale wszystkie ptaki łączy pochodzenie, gdyż każdy z nich jest wyposażony w skrzydła.



Orły, nietoperze i owady mają skrzydła, jednak nie jest to dowód, iż pochodzą one od jednego wspólnego przodka.
Homologia nie stanowi dowodu na słuszność teorii ewolucji. Opiera się ona bowiem jedynie na podobieństwach zewnętrznych wśród gatunków i nie jest naukowo udowodniona. Teza ta przez całe lata, począwszy od połowy XIX wieku, nie została uzasadniona choćby jednym odkryciem naukowym. W żadnej z warstw geologicznych nie natrafiono na fosylia wspólnego przodka organizmów, posiadającego homologiczną strukturę. Także inne argumenty przeczą temu, że homologia może dostarczyć dowodów na słuszność teorii ewolucji:
1. Homologiczne narządy można zaobserwować u zupełnie odmiennych gatunków, pomiędzy którymi ewolucjoniści nie dopatrują się żadnego pokrewieństwa.
2. Kody genetyczne organizmów posiadających podobne narządy są różne.
3. Rozwój embrionalny narządów homologicznych przebiega w odmienny sposób u rozmaitych organizmów.
Pragniemy teraz przyjrzeć się tym argumentom z bliska.
Podobieństwa u ssaków a homologia
Dwa różne gatunki ssaków charakteryzujące się występowaniem dużych zębów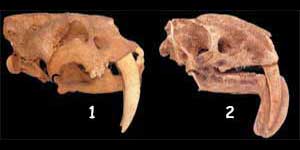
Wręcz bliźniacze podobieństwo pomiędzy należącym do torbaczy wilkiem tazmańskim a będącym przedstawicielem łożyskowców wilkiem północnoamerykańskim jest poważnym problemem dla homologii. Powyżej: czaszki wymienionych zwierząt. Zbieżność między przedstawicielami tych niepokrewnych ewolucyjnie grup sprawia, iż homologiczne hipotezy stają się pozbawione podstaw naukowych.
Wilk północnoamerykański i wilk tazmański


Istnieje duża ilość homologicznych narządów, które stanowią cechy wspólne gatunków, pomiędzy którymi ewolucjoniści nie dopatrują się żadnego pokrewieństwa. Skrzydła występują nie tylko u ptaków, ale także u nietoperzy, które należą do ssaków, u owadów, a nawet u niektórych dinozaurów czyli wymarłych gadów.
Innym przykład to podobieństwa w budowie i funkcjonowaniu oczu różnych organizmów. Człowieka i kałamarnicy nie łączy pokrewieństwo, a jednak budowa i funkcje ich narządów wzroku są bardzo zbliżone. Ten i inne przykłady, które możnaby mnożyć, pokazują, że ewolucjonistyczne stwierdzenie: „występowanie u organizmów organów homologicznych jest dowodem na słuszność teorii ewolucji” nie ma zupełnie podstaw naukowych.
Co więcej, organy te to dla ewolucjonistów kolejna „droga bez wyjścia”. Jeden z uczonych, Frank Salisbury, przyznaje, że występowanie oczu u ekstremalnie zróżnicowanych gatunków jest problemem dla takich jak on zwolenników teorii ewolucji:
„Nawet tak złożona struktura jak oko występuje zarówno u kałamarnicy, jak i u bezkręgowców czy małp wąskonosych. Wystarczająco trudnym zadaniem jest wytłumaczenie, skąd się ono bierze, a myśl, że należy tego dokonywać wiele razy w celu dopasowania się do współczesnej teorii syntetycznej (neodarwinowskiej) przysparza mi bólu głowy.”158
Ewolucjoniści mówią, że wiele organizmów – pomimo podobieństw fizycznych – nie jest ze sobą spokrewnionych. Przykładem mogą być dwie grupy taksonomiczne u ssaków: łożyskowce i torbacze. Zwolennicy teorii ewolucji uważają, że te grupy ssaków oddzieliły się we wcześniejszych epokach i rozwijały niezależnie od siebie. Istnieje jednak w obu grupach znaczna ilość osobników wykazujących wielkie podobieństwa. Amerykańscy biolodzy, Dean Kenyon i Percival Davis, komentują to w następujący sposób:
„Według darwinowskiej teorii, proces ewolucji musiał odbywać się dwa razy: raz dla łożyskowców, a potem – osobno i niezależnie – dla torbaczy. Hipoteza ta doprowadza do zadziwiającej teorii, według której spontaniczny i niekontrolowany proces mutacji oraz selekcji naturalnej w nieznany sposób doprowadził do powstania u niespokrewnionych ze sobą organizmów o takich samych cechach charakterystycznych.”159


Budowa oka kałamarnicy i człowieka jest bardzo podobna. Nie znaczy to jednak, że ludzie i kałamarnice pochodzą od tego samego przodka. Wspólnego rodowodu w tym przypadku nawet ewolucjoniści nie uznają za możliwe.
Owe nieoczekiwane podobieństwa, których ewolucjonistyczni biolodzy nie mogą przedstawić jako przykładów homologii, pokazują, że podobne narządy nie są dowodem potwierdzającym słuszność hipotezy pochodzenia od wspólnego przodka. W takim razie: jakie jest naukowe wytłumaczenie występowania u różnych, w żaden sposób nie spokrewnionych ze sobą grup, organów homologicznych? Odpowiedź na to pytanie jest o wiele starsza niż cała darwinowska teoria. Naukowcy – tacy jak Karol Linneusz czy Richard Owen – wiedzieli, że podobieństwa owe są przykładem i dowodem na wspólny projekt organów różnych zwierząt. Oznacza to, że zbieżności nie powstały w wyniku przekazywania z pokolenia na pokolenie przypadkowo nabytych cech, lecz w rezultacie świadomego planu. Ktoś, kto wiedział, jakie będą funkcje danego organu, zaprojektował go tak, aby funkcje owe mogły być jak najlepiej spełniane.
Aby ewolucjonistyczne postulaty dotyczące homologii miały rację bytu, pomiędzy organizmami powinny występować nie tylko podobieństwa fizyczne, – także ich kody genetyczne musiałyby być homologiczne. W większości przypadków jednak kody te są zróżnicowane; co więcej, jeśli wykazują one podobieństwo, to dotyczy ono często różnych narządów.

Prof. Michael Denton, Evolution: Theory in Crisis
Michael Denton, australijski profesor biochemii, opisuje w swojej książce: Evolution: A Theory in Crisis impas, w jakim znalazła się ewolucjonistyczna interpretacja homologii. Uczony ten pisze: „Homologiczne struktury często powstają na bazie niehomologicznych systemów genetycznych, a sama koncepcja homologii nie ma swego początku w embriologii”.160
Najlepszą ilustracją będzie tu przedstawiany w ewolucjonistycznych źródłach jako dowód, słynny przykład „homologii pięciu palców”. Żyjące na Ziemi bezkręgowce mają po pięć palców u przednich i tylnych odnóży. Mimo iż wygląd tych palców nie jest taki, do jakiego przywykliśmy, organizmy owe zaliczane są na podstawie struktury kości do pentadactyli – pięcioczłonowych. W taki sposób wyposażone są żaby, jaszczurki, wiewiórki czy małpy. Także struktura szkieletu ptaka i nietoperza pasuje do owego ogólnego zarysu.
Ewolucjoniści uważali, że wszystkie te stworzenia mają wspólnego przodka, co przedstawiane było w rozlicznych źródłach biologicznych XX wieku jako dowód na ewolucję. Dopiero odkrycia dokonane w latach osiemdziesiątych na polu genetyki obaliły tę tezę. Okazało się bowiem, że członki owe u różnych grup kontrolowane są przez inne geny. Ewolucjonistyczny biolog, William Fix, opisuje upadek omawianej hipotezy:
„Stare źródła ewolucjonistyczne opierają tezę dotyczącą homologii jedynie na wizualnych podobieństwach w budowie różnych organizmów. Doprowadza to do niebywałych wniosków; na przykład, że ze względu na posiadanie pięciu palców człowiek i ptak pochodzą od jednego wspólnego przodka. Teoria ta miałaby rację bytu, gdybyśmy przyjęli, że owe różne struktury, zmienione dzięki mutacjom i selekcji naturalnej, kierowane były przez tę samą grupę genów. Dzisiaj wiadomo jednak, że tak nie jest; homologiczne organy kontrolowane są przez różne grupy genów. Doprowadziło to do upadku tezy mówiącej, że posiadanie przez organizmy homologicznych członków jest dowodem na wspólne pochodzenie.”161
Innym argumentem podważającym prawdziwość ewolucjonistycznej teorii dotyczącej homologii jest sposób rozwoju embrionalnego gatunków posiadających podobne organy. Rozwój ten powinien przebiegać równolegle, zauważono jednak, że pomiędzy fazami embrionalnymi u organizmów charakteryzujących się homologicznymi narządami występują ogromne różnice.
Krótko mówiąc, odkrycia na polu embriologii i genetyki pokazują, że podobne organy nie świadczą o pochodzeniu od wspólnego przodka. W ten sposób obalona zostaje kolejna z tez Darwina.
Argumenty ewolucjonistów odnoszące się do homologii są bezpodstawne; zarówno jeśli chodzi o narządy, jak i na płaszczyźnie molekularnej. Zwolennicy teorii ewolucji uważają, że kod DNA i struktura białek różnych organizmów jest podobna oraz że te zbieżności są dowodem na ich pochodzenie od jednego wspólnego przodka lub od siebie nawzajem. W rzeczywistości jednak, wyniki porównań na płaszczyźnie molekularnej nie są dla ewolucjonistów optymistyczne. Pomiędzy cząsteczkami organizmów wykazujących podobieństwo fizyczne występują ogromne różnice. Struktura cytochromu C, białka, odgrywającego wielką rolę w procesie oddychania, mocno się różni u przedstawicieli jednego gatunku. W wyniku przeprowadzonych badań okazało się, że rozbieżności między dwoma gatunkami gadów są nawet większe niż pomiędzy niespokrewnionymi gatunkami, np. rybą a ptakiem lub rybą a ssakiem. Zaobserwowano ponadto, że molekularne różnice wśród bakterii, które wydają się bardzo podobne, są często większe niż różnice pomiędzy ssakami a amfibiami czy owadami.162 Badania przeprowadzone na białych i czerwonych cząsteczkach krwi, hormonach i genach doprowadziły do tego samego wniosku. 163 Dr Michael Denton komentuje te odkrycia:
„Na płaszczyźnie molekularnej każda klasa jest specyficzna, odizolowana i w żaden sposób nie powiązana z innymi. Z tego powodu można przyjąć, że podobnie jak fosylia, badania molekularne nie dowodzą słuszności teorii ewolucji – nie wykazują istnienia form przejściowych... Na płaszczyźnie cząsteczkowej żaden organizm nie jest przodkiem innego, nie udowodniono istnienia grup bardziej prymitywnych czy bardziej rozwiniętych... Gdyby wiek temu możliwe było przeprowadzenie takich badań, nie powstałaby teza o ewolucji organicznej.”164
Badania przeprowadzane w latach dziewięćdziesiątych XX wieku pogłębiły jeszcze dylemat ewolucjonistów. Eksperymenty dotyczyły bowiem nie tylko sekwencji białek, jak to bywało do tego czasu, lecz porównywano także łańcuchy r-RNA (rybosomowe RNA). Celem owych doświadczeń było otrzymanie danych do „drzewa ewolucyjnego”. Wyniki eksperymentów nie odpowiadały jednak oczekiwaniom ewolucjonistów. Francuscy biolodzy, Herve Philippe i Patrick Forterre, piszą o tym w jednym z artykułów z 1999 roku:
„Wraz z powiększającą się ilością sekwencji okazywało się, że znaczna ilość filogenów protein zaprzeczała zarówno sobie, jak i drzewu r-RNA”.165
Oprócz wykonywania analiz porównawczych RNA zestawiano także kody genetyczne różnych organizmów; ale i tutaj rezultaty były przeciwne ewolucjonistycznym założeniom „drzewa życia”. Biolodzy molekularni, James Lake, Ravi Jain i Maria Rivera, opisali to w opublikowanym w 1999 roku artykule:
„Naukowcy przeprowadzili analizy porównawcze na wybranych genach różnych organizmów i odkryli, że stosunki pomiędzy nimi przeciwstawiają się ewolucjonistycznemu drzewu życia, wyprowadzonemu tylko z analizy r-RNA.”166
Słuszności teorii filogenii nie dowiodły ani porównania protein, ani r-RNA czy genów. Znany biolog z uniwersytetu w Illinois, Carl Woese, w ten sposób mówi o upadku owej hipotezy:
„Z wyprodukowanych do dziś filogenów protein nie otrzymano żadnego filogenu organizmicznego. Nieścisłości filogenetyczne dało się zauważyć we wszystkich miejscach uniwersalnego drzewa, począwszy od korzeni, aż do gałęzi; wewnątrz i pomiędzy różnymi grupami.”167
Fakt, że porównania cząsteczkowe nie są dowodem za, lecz przeciw teorii ewolucji, podkreślony został także w artykule, który ukazał się w magazynie Science: „Is it Time to Uproot the Tree of Life?” Artykuł ów, autorstwa Elizabeth Pennisi, traktuje o tym, jak nowe badania z dziedziny genetyki porównawczej, które przeprowadzane były przez ewolucjonistycznych biologów i miały na celu wykazanie słuszności teorii ewolucji, doprowadziły do wręcz przeciwnych wniosków. Uczona przekonuje, że świeże dane obalają teorię ewolucji:
„Rok temu przeprowadzono badania na nowopowstałych sekwencjach w genomach znacznej ilości mikroorganizmów, w nadziei, że otrzymane wyniki będą poparciem przyjętych założeń o wcześniejszych epokach życia. Uzyskane informacje nie odpowiadały jednak oczekiwaniom. Porównania genomów nie tylko nie rozjaśniły problemu ewolucyjnego rozwoju głównych grup, ale – wręcz przeciwnie – jeszcze bardziej go zaciemniły. Przy kolejnych ośmiu sekwencjach mikrobicznych sprawa zagmatwała się do końca...
Wielu ewolucjonistycznych biologów wierzyło wprawdzie, że zobaczyli zarys początku życia. Od kiedy łańcuchy DNA umożliwiły porównywanie różnych rodzajów genów, naukowcy oczekiwali, że do drzewa dodadzą jedynie konieczne szczegóły. Jednak „nic nie było dalsze od prawdy”, mówi Claire Fraser, kierownik The Institute for Genomic Research w Rockwill, w stanie Maryland. W wyniku analiz powstało bowiem wiele wersji drzewa życia, które były od poprzedniego odmienne i pozostawały w konflikcie ze sobą.”168
Im więcej przeprowadza się badań i doświadczeń, tym lepiej widać, że koncepcja homologii pozbawiona jest podstaw. Analizy porównawcze protein, r-RNA i genów pokazały, że organizmy uważane przez ewolucjonistów za spokrewnione są w rzeczywistości całkowicie odmienne. W przeprowadzonym w 1996 roku na 88 sekwencjach proteinowych doświadczeniu królik zaliczony został do małp, a nie do gryzoni. Wyniki analizy dokonanej w 1998 roku na 13 genach 19 gatunków zwierząt wykazały przynależność jeżowca do ..., a inne badania przeprowadzone w tym samym roku dały „dowód”, że krowy są bliższe wielorybom niż koniom.169
Jak widać, analizy na płaszczyźnie cząsteczkowej pokazują, że założenia homologii były błędne. Amerykański biolog, Jonathan Wells, streszcza sytuację w ten sposób: „Niedokładności w drzewach pochodzenia i nieoczekiwane wyniki analiz cząsteczkowych sprawiają, że filogenetyka molekularna popada w głęboki kryzys.”170
Kryzys filogenetyki cząsteczkowej oznacza kryzys teorii ewolucji. Nauka po raz kolejny dowiodła, że gatunki nie wywodzą się od jednego wspólnego przodka oraz – że każdy gatunek został stworzony w swojej ostatecznej formie.

Haeckel was an evolutionist even more ardent than Darwin in many respects. For this reason, he did not hesitate to distort the scientific data and devise various forgeries.
To, co kiedyś znane było jako „teoria rekapitulacji”, zostało dawno temu wymazane z literatury naukowej. Mimo to, w niektórych źródłach ewolucjonistycznych teoria rekapitulacji pojawia się jako prawda naukowa. Samo pojęcie „rekapitulacji” to skrót określenia „ontogeny recapitulates phylogeny” (ontogeneza jest powtórzeniem filogenezy), wykreowanego pod koniec XIX wieku przez biologa-ewolucjonistę, Ernsta Haeckla.
Według tego prawa, rozwój osobniczy danego organizmu jest jakby powtórzeniem jego rozwoju rodowego. Zarodek formujący się w łożysku matki podczas rozwoju przyjmuje najpierw cechy ryby, potem gada, a na końcu człowieka.
W późniejszych latach teoria okazała się wyłącznie wytworem wyobraźni. To, co uważano za skrzela – występujące zgodnie z hipotezami teorii rekapitulacji w początkowej fazie rozwoju embrionalnego – okazało się wczesnym etapem rozwoju kanału ucha środkowego. Część embrionu, którą łączono z woreczkiem żółciowym, to w rzeczywistości woreczek, w jakim produkowana była krew dla płodu. Fragment uważany przez Haeckla za ogon był tak naprawdę kręgosłupem, mogącym sprawiać wrażenie ogona, ponieważ pojawiał się, zanim powstały kończyny dolne.
To są uniwersalne prawdy naukowe, nie podważane nawet przez ewolucjonistów. Jeden z założycieli szkoły neodarwinistycznej, George Gaylord Simpson, pisze:
„Haeckel przedstawił rozwój ewolucyjny w błędny sposób. Dzisiaj wiadomo, że ontogeneza nie jest powtórzeniem filogenezy.”171
Magazyn American Scientist podaje, że „teoria rekapitulacji umarła. W latach 50-tych usunięto ją z podręczników szkolnych, ale świat naukowy wykreślił ją znacznie wcześniej, bo już w latach 20-tych.”172
Inny interesujący aspekt rekapitulacji to postać samego jej twórcy, Haeckla, szarlatana fabrykującego niezgodne z prawdą rysunki, na których starał się upodobnić ryby i embriony ludzkie w celu udowodnienia słuszności swej teorii. Kiedy prawda wyszła na jaw, tłumaczył, że także inni ewolucjoniści posługiwali się takimi stworzonymi przez siebie obrazkami:
HAECKEL’S FRAUDULENT DRAWINGS
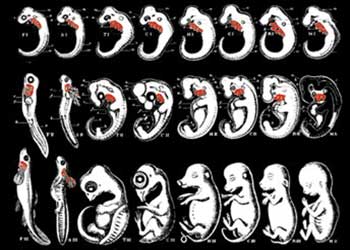
These drawings were fabricated by Haeckel to demonstrate the “similarities” between human and fish embryos. Comparing his sketch with a genuine human embryo, you can see that he has deliberately omitted a large portion of the actual organs. (Francis Hitching, The Neck of the Giraffe: Where Darwin Went Wrong, p. 205)
„Po ujawnieniu, że moje ilustracje były niezgodne z prawdą, powinienem czuć się zawstydzony i zniszczony. Pocieszam się jednak, że na ławie oskarżonych zasiadają wraz ze mną setki tak samo winnych jak ja, a wśród nich wielu uznanych biologów światowej sławy. Większość ilustracji i opracowań z najlepszych nawet podręczników biologii zasługuje na miano fałszerstwa, bo wszystkie one są niedokładne, mniej lub bardziej podretuszowane, schematyczne i skonstruowane.”173
Rzeczywiście, istnieją „setki tak samo winnych, a wśród nich wielu uznanych biologów światowej sławy”, którzy pełni uprzedzeń posługiwali się sfałszowanymi ilustracjami i nieprawdziwymi wynikami doświadczeń w celu obrony teorii, której nie sposób bronić za pomocą naukowych argumentów.
143. Loren Eiseley, The Immense Journey, Vintage Books, 1958, s. 186![]()
144. Charles Darwin, The Origin of Species: A Facsimile of the First Edition, Harvard University Press, 1964, s. 184![]()
145. Norman Macbeth, Darwin Retried: An Appeal to Reason, Harvard Common Press, New York: 1971, s. 33![]()
146. Norman Macbeth, Darwin Retried: An Appeal to Reason, s. 36![]()
147. Loren Eiseley, The Immense Journey, Vintage Books, 1958. s 227![]()
148. Ayrıntılı bilgi için bkz. Jonathan Wells, Icons of Evolution, 2000, s. 159-175![]()
149. Dr. Lee Spetner, "Lee Spetner/Edward Max Dialogue: Continuing an exchange with Dr. Edward E. Max", 2001, http://www.trueorigin.org/spetner2.asp![]()
150. Dr. Lee Spetner, http://www.trueorigin.org/spetner2.asp![]()
151. Dr. Lee Spetner, http://www.trueorigin.org/spetner2.asp![]()
152. Dr. Lee Spetner, http://www.trueorigin.org/spetner2.asp![]()
153. Francisco J. Ayala, "The Mechanisms of Evolution", Scientific American, cilt 239, Eylül 1978, s. 64![]()
154. Dr. Lee Spetner, http://www.trueorigin.org/spetner2.asp![]()
155. S. R. Scadding, "Do 'Vestigial Organs' Provide Evidence for Evolution?", Evolutionary Theory, Cilt 5, Mayıs 1981, s. 173![]()
156. The Merck Manual of Medical Information, Home edition, New Jersey: Merck & Co., Inc. The Merck Publishing Group, Rahway, 1997![]()
157. H. Enoch, Creation and Evolution, New York: 1966, s. 18-19![]()
158. Frank Salisbury, "Doubts About the Modern Synthetic Theory of Evolution", American Biology Teacher, Eylül 1971, s. 338![]()
159. Dean Kenyon & Percival Davis, Of Pandas and People: The Central Question of Biological Origins (Dallas: Haughton Publishing, 1993), p. 33![]()
160. Michael Denton, Evolution: A Theory in Crisis, London, Burnett Books, 1985, s. 145![]()
161. Fix, William, The Bone Peddlers: Selling Evolution (New York: Macmillan Publishing Co., 1984), s. 189![]()
162. W. R. Bird, The Origin of Species Revisited, Thomas Nelson Co., Nashville: 1991, ss. 98-99; Percival Davis, Dean Kenyon, Of Pandas and People, Haughton Publishing Co., 1990, s. 35-38![]()
163. W. R. Bird, The Origin of Species Revisited, s. 98-99, 199-202![]()
164. Michael Denton, Evolution: A Theory in Crisis, London: Burnett Books, 1985, s. 290-91![]()
165. Hervé Philippe and Patrick Forterre, "The Rooting of the Universal Tree of Life is Not Reliable", Journal of Molecular Evolution, vol 49, 1999, p. 510![]()
166. James Lake, Ravi Jain ve Maria Rivera, "Mix and Match in the Tree of Life", Science, vol. 283, 1999, p. 2027![]()
167. Carl Woese, "The Universel Ancestor", Proceedings of the National Academy of Sciences, USA, 95, (1998) p. 6854![]()
168. Elizabeth Pennisi, "MICROBES, IMMUNITY, AND DISEASE: Is It Time to Uproot the Tree of Life?" Science, Volume 284, Number 5418, Issue of 21 May 1999, pp. 1305-1307![]()
169. Jonathan Wells, Icons of Evolution, Regnery Publishing, 2000, p. 51![]()
170. Jonathan Wells, Icons of Evolution, Regnery Publishing, 2000, p. 51![]()
171. G. G. Simpson, W. Beck, An Introduction to Biology, New York, Harcourt Brace and World, 1965, s. 241![]()
172. Keith S. Thompson, "Ontogeny and Phylogeny Recapitulated", American Scientist, Cilt 76, Mayıs / Haziran1988, s. 273![]()
173. Francis Hitching, The Neck of the Giraffe: Where Darwin Went Wrong, New York: Ticknor and Fields 1982, s. 204![]()