La Vera Storia Naturale - II
Uccelli e Mammiferi
Sulla terra ci sono migliaia di specie di uccelli. Ognuna di esse ha caratteristiche distinte. Per esempio, i falchi hanno vista acuta, ali ampie e artigli acuminati, mentre i colibrì, con i loro lunghi becchi, succhiano il nettare dai fiori.
Altri migrano coprendo lunghe distanze fino a specifici posti del mondo. Ma la caratteristica più importante che distingue gli uccelli dagli altri animali è il volo. La maggior parte degli uccelli ha la capacità di volare.
In che modo gli uccelli giunsero all’esistenza? La teoria dell’evoluzione cerca di dare una risposta con un lungo scenario. Secondo questa versione, i rettili sarebbero gli antenati degli uccelli. Circa 150-200 milioni di anni fa, gli uccelli si sarebbero evoluti dai loro antenati rettili. I primi uccelli avrebbero avuto abilità di volo molto scarse. Tuttavia, durante il processo evolutivo, le penne avrebbero sostituito la spessa pelle di questi antichi uccelli, in origine ricoperti di scaglie. Le zampe anteriori si sarebbero coperte completamente di penne e sarebbero diventate ali. Come risultato di una graduale evoluzione, alcuni rettili si sarebbero adattati a volare, diventando gli uccelli di oggi.
Questo scenario viene presentato nelle fonti evolutive come un fatto accertato. Uno studio approfondito dei dettagli dei dati scientifici, però, indica che lo scenario si basa più sulla immaginazione che sulla realtà.
Le origini del volo secondo gli evoluzionisti
Il modo in cui i rettili, come creature abitanti la terra, siano arrivati a volare è un tema che ha suscitato notevoli speculazioni tra gli evoluzionisti. Ci sono due teorie principali. La prima ipotizza che gli antenati degli uccelli siano scesi a terra dagli alberi. Di conseguenza si suppone che questi antenati fossero rettili che vivevano sulla cima degli alberi e che fossero giunti gradualmente a possedere le ali perché saltavano da un ramo all’altro. Questa è nota come la teoria arborea. L’altra, la teoria cursoria (o della “corsa”), suggeriva che gli uccelli fossero passati in aria dalla terra.
Entrambe queste teorie, però, si basano su interpretazioni speculative e non c’è alcuna prova che le supporti. Gli evoluzionisti hanno congetturato una soluzione semplice per questo problema: semplicemente immaginano che la prova esista. Il professor John Ostrom, capo del dipartimento di geologia dell’Università di Yale, che ha proposto la teoria cursoria, spiega questo approccio:
non esiste alcuna evidenza fossile di proavis. È un pre-uccello puramente ipotetico, ma deve essere esistito.78
Questa forma di transizione che la teoria arborea ipotizza che “deve essere vissuta” non è stata mai trovata. La teoria cursoria è ancora più problematica. L’assunto di base della teoria è che le zampe anteriori di alcuni rettili diventarono gradualmente ali perché essi agitavano le zampe per catturare gli insetti. Non viene, però, fornita alcuna spiegazione del modo in cui l’ala, un organo altamente complesso, sia giunta all’esistenza come risultato di questo sbattere.
Un enorme problema per la teoria dell’evoluzione è l’irriducibile complessità delle ali. Solo una struttura perfetta consente alle ali di funzionare, un’ala “sviluppata a metà” non può farlo. In questo contesto, il modello dello “sviluppo graduale” - il meccanismo peculiare postulato dall’evoluzione - non ha senso. Quindi Robert Carroll è costretto ad ammettere che “è difficile spiegare l’evoluzione iniziale delle penne come elementi dell’apparato di volo, perché è difficile vedere in che modo esse potevano funzionare fino a che non avessero raggiunto le grandi dimensioni viste nell‘Archaeopteryx".79 Poi egli ipotizza che le penne potevano essersi evolute per isolamento ma questo non spiega la loro complessa struttura che è specificamente forgiata per volare.

TEORIE IMMAGINARIE, CREATURE IMMAGINARIE
La prima teoria presentata degli evoluzionisti per spiegare le origini del volo sosteneva che i rettili avessero sviluppato le ali mentre cacciavano le mosche (in alto); la seconda teoria fu che diventarono uccelli saltando da un ramo all’altro (sopra). Non c’è, però, alcun fossile di animale che abbia sviluppato le ali gradualmente né alcuna scoperta a dimostrare che una tale cosa fosse persino possibile.
È essenziale che le ali siano strettamente fissate al petto e che posseggano una struttura in grado di sollevare l’uccello e consentirgli di muoversi in tutte le direzioni nonché di restare in aria. È essenziale che ali e penne abbiano una struttura leggera, flessibile e ben proporzionata. A questo punto l’evoluzione è di nuovo in un dilemma. Non risponde alla domanda sul modo in cui questa anatomia perfetta delle ali possa essere venuta fuori come risultato di un accumularsi di mutazioni casuali. Allo stesso modo non offre alcuna spiegazione del modo in cui le zampe anteriori dei rettili siano diventate ali perfette come risultato di un difetto (mutazione) nei geni.
Un’ala formata a metà non può volare. Di conseguenza, anche se presumiamo che la mutazione abbia portato a un piccolo cambiamento nelle zampe anteriori, resta ancora irragionevole ipotizzare che ulteriori mutazioni abbiano contribuito in modo coincidente allo sviluppo di un’ala completa. Questo perché una mutazione delle zampe anteriori non avrebbe prodotto una nuova ala, al contrario, avrebbe solo fatto perdere all’animale le zampe anteriori. Questo lo avrebbe messo in svantaggio rispetto ad altri membri della sua stesa specie. Secondo le regole della teoria dell’evoluzione, la selezione naturale avrebbe presto eliminato questa creatura difettosa.
Secondo la ricerca biofisica, le mutazioni sono modifiche che avvengono molto raramente. Di conseguenza è impossibile che un animale inabile abbia aspettato milioni di anni perché le sue ali si sviluppassero completamente attraverso leggere mutazioni, soprattutto quando queste mutazioni hanno effetti dannosi nel tempo.
Uccelli e dinosauri
La teoria dell’evoluzione sostiene che gli uccelli si siano evoluti da teropodi carnivori e bipedi. Un confronto tra uccelli e rettili, però, rivela che i due hanno caratteristiche molto distinte che rende improbabile che l’uno si sia evoluto nell’altro.
Ci sono varie differenze strutturali tra uccelli e rettili, una delle quali riguarda la struttura ossea. A causa della loro mole, i dinosauri – gli antenati degli uccelli secondo gli evoluzionisti – avevano ossa spesse e compatte. Gli uccelli, al contrario, sia viventi che estinti, hanno ossa cave che sono molto leggere come devono essere perchè avvenga il volo.
Un’altra differenza tra rettili e uccelli è la loro struttura metabolica. I rettili hanno la struttura metabolica più lenta del regno animale (l’affermazione secondo cui i dinosauri avrebbero avuto un metabolismo veloce a sangue caldo resta una speculazione). Gli uccelli, d’altro canto, sono all’estremo opposto dello spettro metabolico. Per esempio, la temperatura corporea di un passero può arrivare fino a 48°C a causa del suo metabolismo veloce. I rettili, d’altro canto, non hanno la capacità di regolare la propria temperatura corporea. Per scaldarsi, espongono il corpo alla luce del sole. In parole povere, i rettili sono gli animali che consumano meno energia e gli uccelli quelli che ne consumano di più.
Uno dei più noti ornitologi del mondo, Alan Feduccia, dell’Università della North Carolina, si oppone alla teoria secondo cui gli uccelli sarebbero parenti dei dinosauri, pur essendo egli stesso un evoluzionista. Rispetto allo scenario rettili-uccelli, Feduccia ha da dire quanto segue:
Ebbene, io studio i crani degli uccelli da 25 anni e non vedo alcuna somiglianza di sorta. Semplicemente non la vedo… Le origini teropodi degli uccelli, secondo me, saranno il più grande imbarazzo della paleontologia del ventesimo secolo. 80
Anche Larry Martin, specialista di uccelli antichi dell’Università del Kansas, si oppone alla teoria secondo cui gli uccelli discendono dai dinosauri. Discutendo le contraddizioni in cui l’evoluzione cade su questo argomento, egli afferma:
a dire la verità, se dovessi sostenere l'origine degli uccelli dai dinosauri con quelle caratteristiche, sarei imbarazzato ogni volta che dovessi alzarmi e parlarne.81
Tuttavia, nonostante tutte le scoperte scientifiche, lo scenario infondato della “evoluzione dinosauro-uccello” è ancora sostenuto con insistenza. Pubblicazioni popolari amano particolarmente questo scenario. Allo stesso tempo, concetti che non forniscono alcun supporto per lo scenario stesso sono presentati come prove per l’immaginaria “evoluzione dinosauro-uccello”.
In alcune pubblicazioni evoluzioniste, per esempio, si pone l’accento sulle differenze tra le ossa delle anche dei dinosauri per supportare la tesi secondo cui gli uccelli sarebbero discesi da loro. Queste cosiddette differenze esistono tra dinosauri classificati come saurischiani (simili a rettili, specie con cingolo dell’anca) e ornitischi (simili ad uccelli, specie con cingolo dell’anca). Questo concetto di dinosauri con cingoli dell’anca simili a quelli degli uccelli viene adoperato di tanto in tanto quale prova dell’ipotetico collegamento dinosauro-uccello. La differenza nel cingolo dell’anca, comunque, non è affatto una prova che gli uccelli si siano evoluti dai dinosauri. Questo perché i dinosauri ornitischi non somigliano agli uccelli per quanto riguarda altre caratteristiche anatomiche. Per esempio l’Anchilosauro è un dinosauro classificato come ornitischio, con zampe corte, un corpo gigantesco e pelle coperta da scaglie che somigliavano ad una corazza. D’altro canto lo Struziomimo, che somiglia agli uccelli in alcune strutture anatomiche (zampe posteriori lunghe, zampe anteriori corte e struttura sottile), è in effetti un saurischiano.82
In breve, la struttura del cingolo dell’anca non è una prova della relazione evolutiva tra uccelli e dinosauri. Dire che i dinosauri somigliano agli uccelli perché i loro cingoli d’anca sono simili, vuol dire ignorare altre significative differenze anatomiche tra le due specie che rendono il collegamento evolutivo tra di loro insostenibile da punto di vista evoluzionista.
La struttura peculiare dei polmoni degli uccelli
Un altro fattore che dimostra l’impossibilità dello scenario evolutivo rettili-uccelli è la struttura dei polmoni di questi ultimi che non possono essere spiegati con l’evoluzione.
Nelle creature che vivono sulla terra il flusso d’aria è bidirezionale. Al momento dell’inalazione, l’aria viaggia attraverso i passaggi nei polmoni (i tubi bronchiali) finendo nelle minuscole sacche aeree (alveoli). Lo scambio di ossigeno e di biossido di carbonio ha luogo lì. Poi, al momento dell’espirazione, l’aria usata torna indietro e trova la strada fuori dai polmoni con lo stesso percorso.
L’ESCLUSIVO SISTEMA SCHELETRICO DEGLI UCCELLI
A differenza delle ossa di dinosauri e rettili, le ossa degli uccelli sono cave. Questo dà al corpo stabilità e leggerezza, La struttura dello scheletro degli uccelli si utilizza per progettare aerei, ponti e strutture moderne.

Le ossa dei dinosauri sono spesse e compatte a causa della loro struttura massiccia mentre le ossa di uccelli viventi ed estinti sono cave e quindi molto leggere.
Negli uccelli, invece, l’aria è unidirezionale. L’aria nuova entra da un’estremità e quella usata esce dall’altra. Grazie a speciali sacche d’aria collocate lungo tutti i passaggi, l’aria fluisce sempre in una sola direzione attraverso i polmoni. In tal modo, gli uccelli sono in grado di prendere aria senza interruzione. Questo soddisfa i bisogni di energia degli uccelli. Questo sistema respiratorio altamente specializzato è spiegato da Michael Denton nel suo libro A Theory in Crisis:
Nel caso degli uccelli, i bronchi più importanti si dividono in minuscoli tubi che permeano il tessuto polmonare. I cosiddetti parabronchi alla fine si uniscono formando un vero sistema circolatorio in modo che l’aria fluisca in un’unica direzione attraverso i polmoni. .. La struttura dei polmoni negli uccelli e il funzionamento complessivo del sistema respiratorio è del tutto peculiare. Non si conosce alcun altro polmone in altre specie di vertebrati che si avvicini al sistema degli uccelli. Inoltre è identico in tutti i dettagli essenziali in uccelli diversi come i colibrì, gli struzzi e i falchi. 83
La cosa importante è che i polmoni dei rettili, con il loro flusso d’aria bidirezionale, non si sarebbero potuti evolvere diventando i polmoni degli uccelli, con il loro flusso unidirezionale, perché non è possibile un modello intermedio tra di loro. Perché una creatura viva, deve continuare a respirare, e un rovesciamento della struttura dei polmoni con una modifica del modello stesso avrebbe come fine inevitabile la morte. Secondo l’evoluzione, questa modifica deve essere avvenuta gradualmente nel corso di milioni di anni mentre una creatura i cui polmoni non funzionano muore entro pochi minuti.
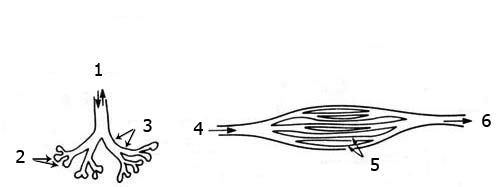
1. flusso aereo
2. alveoli
3. bronchi
4. l’aria entra
5. parabronchi
6. l’aria esce
I polmoni degli uccelli funzionano in un modo contrario rispetto a quelli degli animali di terra. Questi ultimi inalano ed esalano dagli stessi passaggi. Al contrario, negli uccelli l’aria passa in continuazione attraverso i polmoni in una sola direzione. Questo è reso possibile da speciali sacche aeree presenti in tutto il polmone. Grazie a questo sistema, i cui dettagli possono essere visti sopra, gli uccelli respirano in continuazione. Questo sistema è peculiare degli uccelli che hanno bisogno di alti livelli di ossigeno durante il volo. È impossibile che questa struttura si sia evoluta nei polmoni dei rettili, perché qualsiasi creatura con una forma “intermedia” tra i due tipi di polmoni non sarebbe stata in grado si respirare.
Michael Denton afferma che è impossibile dare una spiegazione evolutiva ai polmoni degli uccelli.
E’ assolutamente difficile da immaginare come un sistema respiratorio estremamente diverso potrebbe essersi evoluto gradualmente dal modello generale dei vertebrati, specialmente tenendo presente che il mantenimento delle funzioni respiratorie è assolutamente fondamentale per la vita di un organismo nella misura in cui il più piccolo malfunzionamento porta alla morte in pochi minuti. Proprio come le piume non possono funzionare quali organi di volo fino a che i ganci e le barbule non si adattano in modo da completarsi perfettamente tra loro, così i polmoni degli uccelli non possono funzionare come organi della respirazione fino a che il sistema parabronchiale che li permea e il sistema di sacche aeree che garantisce ai parabronchi il loro apporto di aria non siano entrambi pienamente sviluppati e in grado di funzionare assieme in una maniera perfettamente integrata. 84
In breve, il passaggio da un polmone terrestre a quello degli uccelli è impossibile, perché una forma intermedia non avrebbe alcuna utilità.
Un altro punto che deve essere citato qui è che i rettili hanno un sistema respiratorio a diaframma, mentre gli uccelli hanno un sistema di sacche aeree addominali. Anche queste strutture diverse rendono impossibile qualsiasi evoluzione tra i due tipi di polmone, come osserva nel passaggio che segue John Ruben, un’autorità riconosciuta nel campo della fisiologia respiratoria:
Le prime fasi nel passaggio da un antenato con ventilazione a diaframma al sistema a sacche aeree addominali degli uccelli, avrebbero avuto bisogno di una selezione per un’ernia diaframmatica nei taxa di transizione tra teropodi e uccelli. Una condizione così debilitante avrebbe immediatamente compromesso l’intero apparato ventilatorio polmonare e sembra improbabile che sia stato di alcun vantaggio selettivo. 85
Un’altra interessante caratteristica strutturale dei polmoni degli uccelli che sfida l’evoluzione è che non sono mai privi di aria, e quindi mai in pericolo di collasso. Michael Denton spiega la posizione:
Proprio il modo in cui un sistema respiratorio diverso potrebbe essersi evoluto gradualmente dal modello generale dei vertebrati senza alcun tipo di direzione è, ancora una volta, molto difficile da immaginare, specialmente tenendo presente il fatto che il mantenimento delle funzioni respiratorie è assolutamente fondamentale per la vita di un organismo. Inoltre, la funzione e la forma uniche dei polmoni degli uccelli richiedono numerosi ulteriori adattamenti unici durante lo sviluppo degli uccelli stessi. Come spiega H. R. Dunker, una delle autorità mondiali in questo campo, poiché per prima cosa i polmoni degli uccelli sono fissati rigidamente alla parete del corpo e non possono quindi espandersi in volume e, in secondo luogo, a causa del diametri ridotto dei capillari polmonari e della risultante alta tensione superficiale di qualsiasi liquido all’interno di essi, i polmoni degli uccelli non potrebbero gonfiarsi partendo da uno stato collassato come accade a tutti gli altri vertebrati dopo la nascita. I capillari aerei non collassano mai, come fanno gli alveoli di altre specie di vertebrati; piuttosto, poiché crescono nel tessuto polmonare, i parabronchi sono sin dall’inizio tubi aperti pieni o di aria o di liquido. 86
In altre parole, i passaggi nei polmoni degli uccelli sono tanto stretti che le sacche aeree all’interno non possono riempirsi di aria e poi svuotarsi, come nel caso delle creature che vivono sulla terra.
Se il polmone di un uccello si sgonfiasse completamente, l’uccello non sarebbe mai in grado di rigonfiarlo oppure, almeno, avrebbe grandi difficoltà a farlo. Per questo motivo, le sacche aeree situate lungo tutto il polmone consentono un costante passaggio di aria, proteggendo così i polmoni dallo sgonfiamento.
Naturalmente questo sistema, che è completamente diverso dai polmoni dei rettili e di altri vertebrati ed è basato sul più delicato equilibrio, non può essere comparso con mutazioni inconsapevoli, fase per fase, come sostiene l’evoluzione. Ecco in che modo Denton descrive questa struttura del polmone degli uccelli, che ancora invalida il darwinismo:
I polmoni degli uccelli ci portano molto vicino a rispondere alla sfida di Darwin: "se si potesse dimostrare l'esistenza di un qualsiasi organo complesso che non si sia potuto essere formato attraverso modificazioni numerose, successive, lievi, la mia teoria dovrebbe assolutamente cadere."87
Piume degli uccelli e scaglie dei rettili
Un altro invalicabile scoglio tra uccelli e rettili è rappresentato dalle penne, che sono peculiari degli uccelli. I corpi dei rettili sono coperti di scaglie e quelli degli uccelli di penne. L’ipotesi secondo cui le penne degli uccelli si svilupparono dalle scaglie dei rettili è completamente infondata ed è in realtà confutata dai reperti fossili, come ammette la paleontologa evoluzionista Barbara Stahl:
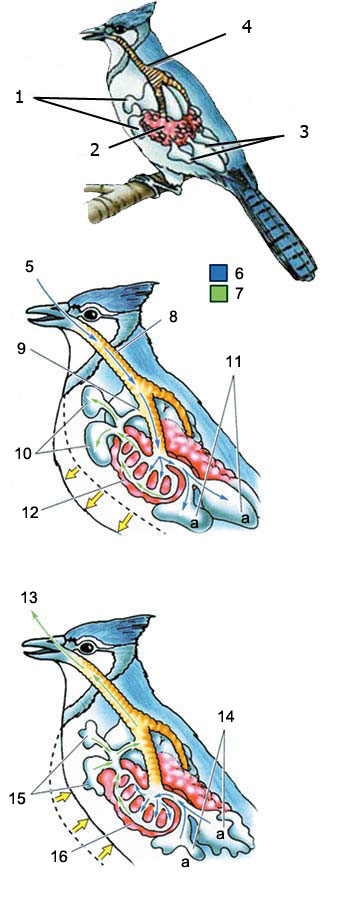
13. Aria esausta
14. L’aria fresca esce dalle sacche aeree posteriori verso i polmoni.
15. L’aria esausta è espulsa attraverso le sacche aeree anteriori.
16. Polmone
a: Air pur
LO SPECIALE SISTEMA RESPIRATORIO DEGLI UCCELLI
INSPIRAZIONE: L’aria che entra nei passaggi respiratori degli uccelli va verso il polmone e verso le sacche aeree dietro di essi. L’aria usata viene trasferita nelle sacche aeree anteriori.
ESPIRAZIONE:Quando l’uccello espira, l’aria fresca delle sacche aeree posteriori entra nei polmoni. Con questo sistema, l’uccello è in grado di usufruire di una costante riserva di aria fresca nei polmoni.
Il y a beaucoup de détails dans ce système pulmonaire, qui sont présentés de façon très simplifiée dans ces diagrammes. Il existe, par exemple, des valves particulières où les sacs rejoignent les poumons, qui permettent à l'air de circuler dans la bonne direction. Tout cela montre clairement que la "création" est ici à l'oeuvre. Ces systèmes spéciaux assènent non seulement un coup décisif à la théorie de l'évolution, mais ils sont aussi parmi les nombreuses preuves de la réalité de la création.
1. sacche aeree anteriori
2. polmone
3. sacche aeree posteriori
4. trachea
5. Aria fresca
6. Aria fresca
7. Aria esausta
8. Trachea
9. L’aria fresca nonpassa attraverso le sacche aeree anteriori
10. Le sacche aeree anteriori si riempiono dell’aria esausta che viene dai polmoni
11. Le sacche aeree posteriori si riempiono di aria fresca
12. Polmone
In che modo [le penne] comparvero inizialmente, presumibilmente dalle scaglie dei rettili, sfida l'analisi... Sembra, dalla complessa costruzione delle penne, che la loro evoluzione dalle scaglie dei rettili, avrebbe richiesto un periodo di tempo immenso e comportato una serie di strutture intermedie. Finora, i reperti fossili non supportano tale ipotesi. 88
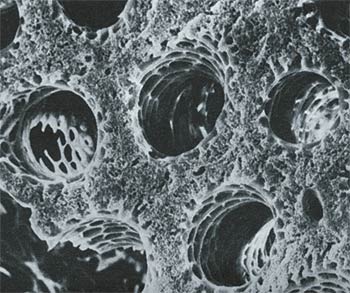
I tubi parabronchiali che consentono all’aria di circolare nella giusta direzione nei polmoni degli uccelli. Ciascun tubo ha un diametro di soli 0,5 mm.
A.H. Brush, professore di fisiologia e neurobiologia dell'Università del Connecticut, accetta questa realtà anche se è egli stesso un evoluzionista. “Ogni caratteristica, dalla struttura e l’organizzazione del gene fino allo sviluppo, la morfogenesi e l’organizzazione dei tessuti, è diversa (tra piume e scaglie)”. 89 Inoltre il professor Brush esamina la struttura delle proteine delle piume degli uccelli e sostiene che è “unica tra i vertebrati”.90
Non c’è alcuna prova fossile a dimostrare che le piume degli uccelli si siano evolute dalle scaglie dei rettili. Al contrario, le penne appaiono all’improvviso nei reperti fossili, osserva il prof. Brush, come un carattere “innegabilmente unico” che distingue gli uccelli”.91 Inoltre, nei rettili non è stata rilevata ancora alcuna struttura epidermica che possa dare origine alle piume degli uccelli.92
Molti fossili sono stati finora oggetto di speculazioni su “dinosauri con le penne”, ma uno studio dettagliato le ha sempre confutate. L’importante ornitologo Alan Feduccia scrive quanto segue in un articolo dal titolo "On Why Dinosaurs Lacked Feathers":
Le penne sono caratteristiche uniche degli uccelli e non c’è alcuna struttura intermedia nota tra le scaglie dei rettili e le penne degli uccelli.Nonostante le speculazioni sulla natura delle scaglie allungate trovate in forme come il Longisquama ... ipotizzate come strutture simili alle penne, semplicemente non c’è alcuna prova dimostrabile che in realtà lo siano.93
Segnali della creazione nelle penne
D’altro canto le penne degli uccelli hanno una struttura così complessa che il fenomeno non può essere spiegato con processi evolutivi. Come ben sappiamo, c’è un’asta che corre lungo il centro della penna. Collegate all’asta ci sono le pinnule. La pinnula è costituita da piccoli fili chiamati barbe. Queste barbe, di diversa lunghezza e rigidità, sono ciò che dà all’uccello la sua natura aerodinamica. Ma la cosa più interessante è che ciascuna barba ha migliaia di fili ancora più piccoli fissati ad essa, chiamati barbule. Le barbule sono collegate alle barbicelle, con minuscoli ganci microscopici, chiamati amuli. Ciascun filo è agganciato a un altro opposto, come i ganci di una cerniera.
Solo la penna di sollevamento ha circa 650 barbe su ciascun lato dell’asta. Circa 600 barbule si ramificano dalle barbe. Ciascuna di queste barbule è tenuta insieme da 390 gancetti. I gancetti si incastrano come i denti di entrambi i lati di una cerniera. Se, per qualunque motivo, i gancetti si aprono, l’uccello può facilmente riportare le penne alla forma originaria scuotendosi o lisciandosi le penne con il becco.
Affermare che la complessa struttura delle penne potrebbe essere emersa dall’evoluzione delle scaglie dei rettili attraverso mutazioni casuali è semplicemente una credenza dogmatica, senza alcun fondamento scientifico. Persino uno dei decani del darwinismo, Ernst Mayr, ha fatto questa confessione sull’argomento alcuni anni fa:
Ci vuole una notevole dose di fode per ritenere che sistemi finemente equilibrati come certi organi di senso (l’occhio dei vertebrati o le penne degli uccelli) potrebbero essere migliorati attraverso mutazioni casuali.94
Darwin si sentì costretto a prendere in considerazione le penne. L’estetica perfetta delle piume del pavone lo aveva fatto stare “male” (secondo le sue stesse parole). In una lettera scritta ad Asa Gray il 3 aprile 1860, egli disse: “ricordo bene il tempo in cui il pensiero dell’occhio mi fece gelare, ma ho superato questa fase…” E poi continuava: "... e ora insignificanti particolari della struttura spesso mi fanno sentire a disagio. La vista di una piuma nella coda di un pavone, ogni volta che la guardo, mi fa star male!”95
In breve, le enormi differenze strutturali tra le penne degli uccelli e le scaglie dei rettili e la struttura incredibilmente complessa delle penne, dimostrano chiaramente l’infondatezza dell’affermazione secondo cui le penne si evolvettero dalle scaglie.

Il fossile di Sinosauropteryx, che i paleontologi evoluzionisti annunciarono essere un "dinosauro pennuto", ma che, successivamente, si rivelò non esserlo.

SCAGLIE DEI RETTILI
Le scaglie che coprono il corpo dei rettili sono totalmente diverse dalle penne degli uccelli. A differenza delle penne, le scaglie non si estendono sotto la pelle ma sono solo uno strato duro sulla superficie del corpo dell’animale. Geneticamente, biochimicamente e anatomicamente, le scaglie non hanno alcuna somiglianza con le penne. Questa grande differenza tra di loro dimostra ancora una volta che lo scenario dell’evoluzione da rettili a uccelli è infondato.
Il malinteso dell’Archaeopteryx
In risposta alla domanda se ci sia qualche evidenza fossile della “evoluzione rettile-uccello”, si pronuncia il nome di una sola creatura. È il fossile di un uccello chiamato Archaeopteryx, una delle più conosciute tra le cosiddette forme di transizione che pochissimi evoluzionisti ancora difendono.
L’Archaeopteryx, il cosiddetto antenato degli uccelli dei nostri giorni, secondo gli evoluzionisti, visse circa 150 milioni di anni fa. La teoria sostiene che alcuni piccoli dinosauri, come i Velociraptor o i Dromeosauri, si sarebbero evoluti acquisendo le ali e poi cominciando a volare. Si presume, quindi, chel’Archaeopteryx sia una forma di transizione che derivava dai suoi antenati dinosauri e che aveva cominciato a volare per la prima volta.
Studi più recenti dei fossili di Archaeopteryx, però, indicano che questa spiegazione manca di qualsiasi fondamento scientifico. Non è assolutamente una forma di transizione, ma una specie estinta di uccelli che aveva alcune insignificanti differenze rispetto agli uccelli moderni.
LA COMPLESSA STRUTTURA DELLE PENNE DEGLI UCCELLI

Quando si studiano da vicino le penne degli uccelli, emerge una struttura molto delicata. Sono più sottili dei capelli e hanno speciali ganci che consentono loro di fissarsi tra di loro. Le immagini mostrano penne di uccelli allargate progressivamente.

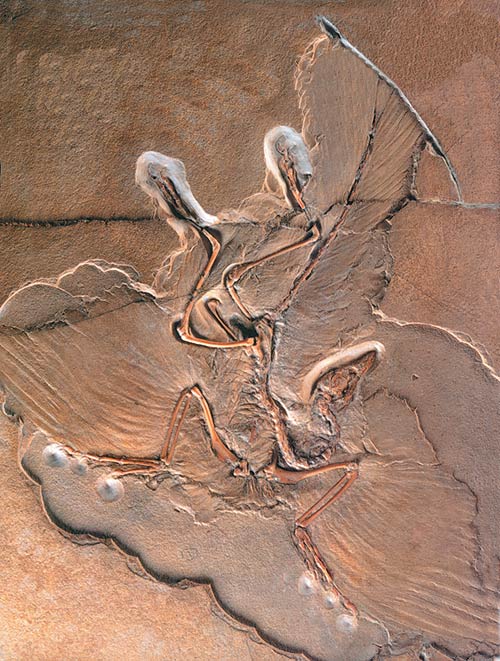
Una delle prove importanti che l’Archaeopteryx era un uccello volante è la struttura asimmetrica delle sue penne. Sopra una delle penne fossili della creatura.
La tesi secondo cui l’Archaeopteryx sarebbe stato un "semi-uccello" che non poteva volare perfettamente, era popolare nei circoli evoluzionisti fino a non molto tempo fa. L’assenza di uno sterno in questa creatura era ritenuta come la prova più importante che questo uccello non potesse volare correttamente (lo sterno è un osso che si trova sotto il torace a cui sono fissati i muscoli necessari per il volo. Oggi, questo osso si osserva in tutti gli uccelli volanti e non volanti, e anche nei pipistrelli, mammiferi volanti che appartengono a una famiglia molto diversa). Il settimo fossile di Archaeopteryx,trovato nel 1992, confutò questa argomentazione. Infatti, in questo fossile scoperto di recente, si scoprì che lo sterno, che per lungo tempo era stato ritenuto mancante dagli evoluzionisti, in realtà era presente. Questo fossile fu descritto nel giornale Nature come segue:
Il settimo esemplare di Archaeopteryx scoperto di recente conserva uno sterno rettangolare parziale, a lungo ipotizzato, mai documentato prima. Questo sterno è una prova dei forti muscoli del volo, ma la capacità di intraprendere lunghi voli è dubbia. 96
Questa scoperta invalida il principale sostegno delle affermazioni secondo cui l'Archaeopteryx sarebbe stato un semi-uccello incapace di volare correttamente.
Inoltre, la struttura delle penne dell’uccello divenne una delle prove più importanti a conferma del fatto che l’Archaeopteryxera un uccello volante nel vero senso della parola. La struttura asimmetrica delle penne dell’Archaeopteryx non si distingue da quella degli uccelli di oggi e indica che poteva volare perfettamente. Come afferma l’eminente paleontologo Carl O. Dunbar: "In ragione delle sue ali, [l’Archaeopteryx deve essere] chiaramente classificato come un uccello”. 97 Il paleontologo Robert Carroll spiega ulteriormente questo argomento:
La geometria delle penne del volo dell’Archaeopteryx è identica a quella degli uccelli volanti moderni, laddove gli uccelli non volanti hanno penne simmetriche. Il modo in cui le penne sono disposte sull’ala ricade anch’esso nell'ambito degli uccelli moderni… Secondo Van Tyne e Berger, la dimensione e la forma relative delle ali dell'Archaeopteryx sono simili a quelle degli uccelli che si muovono attraverso aperture limitate nella vegetazione, come i gallinacei, le colombe, le beccacce, i picchi e molti passeracei…Le penne del volo sono in stasi da almeno 150 milioni di anni…98
Un altro fattore rivelato dalla struttura delle penne dell'Archaeopteryx fu il suo metabolismo a sangue caldo tipico degli uccelli. Come discusso in precedenza, i rettili e i dinosauri sono animali a sangue freddo il cui calore corporeo fluttua con la temperatura ambiente, piuttosto che essere regolato omeostaticamente. Una funzione molto importante delle penne degli uccelli è il mantenimento di una temperatura corporea costante. Il fatto che l'Archaeopteryx avesse penne, dimostra che era un vero uccello a sangue caldo, che aveva bisogno di conservare il calore corporeo, a differenza dei dinosauri.
I denti e gli artigli dell'Archaeopteryx
Due punti importanti sui quali i biologi evoluzionisti fanno affidamento quando affermano che l'Archaeopteryx era una forma di transizione, sono gli artigli sulle ali e i denti.
È vero che l’Archaeopteryx aveva artigli sulle ali e denti in bocca ma queste caratteristiche non implicano che la creatura avesse alcuna relazione con i rettili. Inoltre, due specie di uccelli oggi viventi, il turaco e l'hoatzin, hanno artigli per aggrapparsi ai rami. Queste creature sono completamente uccelli, senza alcuna caratteristica dei rettili. Ecco perché è del tutto infondato sostenere che l'Archaeopteryx sia una forma di transizione soltanto per la presenza degli artigli sulle ali.
Il più noto Archaeopteryx fossile, in mostra a Berlino.
Neppure i denti nel becco dell’Archaeopteryx implicano che sia stato una forma di transizione. Gli evoluzionisti sbagliano nel dire che i denti siano una caratteristica dei rettili, poiché non sono tipici dei rettili. Oggi alcuni rettili hanno i denti, mentre altri no. Inoltre, l'Archaeopteryx non è la sola specie di uccelli che presenti i denti. È vero che ci sono uccelli privi di denti oggi, tuttavia se guardiano i reperti fossili, vediamo che sia ai tempi dell'Archaeopteryx che in seguito e anche fino a tempi abbastanza recenti, esisteva un distinto genere di uccelli che può essere classificato come "uccello provvisto di denti".
Il punto più importante, tuttavia, è che la struttura dentale dell'Archaeopteryx e di altri uccelli con denti è del tutto diversa da quella dei loro supposti antenati, i dinosauri. I famosi ornitologi L.D.Martin, J.D. Stewart e K.N. Whetstone osservarono che gli Archaeopteryx e altri uccelli simili, avevano denti non seghettati con basi strette e radici espanse. Eppure i denti dei dinosauri teropodi, i presunti antenati di questi uccelli, erano seghettati, con radici strette. 99 Questi ricercatori paragonarono anche le ossa della caviglia dell'Archaeopteryxa quelle dei loro presunti antenati, i dinosauri e non osservarono alcuna somiglianza. 100
Gli studi di anatomisti quali S. Tarsitano, M.K. Hecht e A.D. Walker rivelarono che alcune delle somiglianze, riscontrate da John Ostrom e altri, tra le membra dell'Archaeopteryx e i dinosauri, erano in realtà interpretazioni errate.101 Per esempio A.D. Walker ha analizzato la regione dell’orecchio dell’Archaeopteryx ed ha scoperto che è identica a quella degli uccelli dei giorni nostri. 102
Inoltre, J. Richard Hinchliffe, dell’Istituto di Scienze Biologiche dell’Università del Galles, ha studiato l’anatomia degli uccelli e dei loro presunti antenati rettili, usando le moderne tecniche all’isotopo, e ha scoperto che le tre dita delle zampe anteriori dei dinosauri sono I-II-III, mentre le dita delle ali degli uccelli sono II-III-IV. Questo crea un grave problema ai sostenitori del collegamento Archaeopteryx-dinosauro.103 Hinchliffe ha pubblicato i suoi studi e le sue osservazioni in Science nel 1997, in cui scrisse:
I dubbi sull’omologia tra le dita dei teropodi e quelle degli uccelli ci ricordano alcuni dei problemi nelle ipotesi sulla “origine da dinosauri”. Tra questi, quelli che seguono: (i) La zampa anteriore molto più piccola del teropode (in relazione alla dimensione del corpo) se confrontata all’ala dell’Archaeopteryx. Tali piccole membra non sono convincenti come protoali, capaci di permettere di spiccare il volo da terra ai relativamente pesanti dinosauri. (ii) La rarità nei teropodi dell’osso del polso semiluminato, noto solo in quattro specie (tra cui il Deinonychus). La maggior parte dei teropodi ha elementi del polso relativamente grandi, difficili da considerare omologhi con quelli dell'Archaeopteryx. (iii) Il paradosso temporale; infatti, la maggior parte dei dinosauri teropodi, e in particolare il dromeosauro simile agli uccelli, appaiono tutti molto più tardi nei reperti fossili rispetto all’Archaeopteryx.
Come nota Hinchliffe, il “paradosso temporale” è uno dei fatti che infliggono un colpo fatale alle ipotesi evoluzioniste sull’Archaeopteryx. Nel suo libro Icons of Evolution, il biologo americano Jonathan Wells nota che l’Archaeopteryx è diventato “un’icona” della teoria dell’evoluzione, mentre le prove mostrano chiaramente che questa creatura non è l’antenato primitivo degli uccelli. Secondo Wells, una delle indicazioni di ciò è che i dinosauri teropodi — i presunti antenati dell’Archaeopteryx— sono in realtà successivi all’Archaeopteryx: "Rettili a due zampe che correvano sul terreno, e con altre caratteristiche che ci si poteva aspettare in un antenato dell’Archaeopteryx, comparvero successivamente."104
Tutte queste scoperte indicano che l’Archaeopteryxnon era un anello di transizione, ma solo un uccello che ricadeva in quella categoria che può essere chiamata degli “uccelli dentati”. Collegare questa creatura ai dinosauri teropodi non ha alcuna validità. In un articolo dal titolo "The Demise of the 'Birds Are Dinosaurs' Theory," il biologo americano Richard L. Deem scrive quanto segue circa l’Archaeopteryxe l’affermazione dell’evoluzione uccello-dinosauro:
I risultati degli studi recenti dimostrano che le zampe dei dinosauri teropodi derivano dalle dita I, II e III, laddove le ali degli uccelli, anche se sembrano simili in termini di struttura, derivano dalle dita II, III e IV... Ci sono altri problemi nella teoria che afferma che “gli uccelli sono dinosauri”. La zampa anteriore del teropode è molto più piccola (in relazione alla dimensione del corpo) di quella dell’Archaeopteryx. La piccola “protoala” del teropode non convince molto, specialmente considerando il peso piuttosto notevole di questi dinosauri. La grande maggioranza dei teropodi manca di osso del polso semiluminato e ha un gran numero di altri elementi del polso che non hanno alcuna omologia con le ossa dell’Archaeopteryx. Inoltre, in quasi tutti i teropodi, il nervo V1 esce dalla scatola cranica lateralmente, insieme con altri nervi, mentre negli uccelli esce dalla parte anteriore della scatola cranica, attraverso l’apposito foro. C’è anche un problema meno importante, cioè che la grande maggioranza dei teropodi comparve dopo l’Archaeopteryx. 105
L'Archaeopteryx e gli altri fossili di antichi uccelli

Come per l‘Archaeopteryx, ci sono unghie simili ad artigli sulle ali dell’uccello Opisthocomus hoazin, che vive ai giorni nostri
Anche alcuni fossili recentemente rinvenuti hanno invalidato per altri versi lo scenario evoluzionista a proposito dell’Archaeopteryx.
Lianhai Hou e Zhonghe Zhou, due paleontologi dell'Istituto Cinese di Paleontologia dei Vertebrati, hanno scoperto un nuovo uccello fossile nel 1995, denominato Confuciusornis. Questo uccello risale approssimativamente allo stesso periodo dell'Archaeopteryx (circa 140 milioni di anni fa), ma non è fornito di alcun tipo di dente. Il suo becco e le sue ali, inoltre, sono del tutto simili a quelle degli uccelli attuali. Il Confuciusornis ha la medesima struttura scheletrica degli uccelli attuali, tuttavia presenta artigli sulle ali, proprio come l'Archaeopteryx. Un’altra struttura peculiare degli uccelli, detta "pigostilo", che sostiene le penne timoniere, era presente nel Confuciusornis.106 In breve, questo fossile, contemporaneo dell'Archaeopteryx, considerato in precedenza il più antico antenato di tutti gli uccelli e accettato come un semi-rettile, ha un'enorme somiglianza con i volatili moderni. Questo fatto inficia tutte le tesi evoluzioniste che reputavano l'Archaeopteryx il primitivo antenato di tutti gli uccelli.
Un altro fossile rinvenuto in Cina ha creato una confusione addirittura maggiore. Nel novembre 1996 l'esistenza di un uccello, risalente a 130 milioni di anni fa, detto Liaoningornis, fu annunciata su Science da L. Hou, L. D. Martin e Alan Feduccia. Il Liaoningornis presentava uno sterno sul quale si innestavano i muscoli del volo, del tutto simile a quello degli uccelli attuali.107 Questo uccello era difficilmente distinguibile dagli uccelli attuali anche per altri motivi. La sola differenza era rappresentata dai denti nel becco. Questo fatto mostrò che gli uccelli forniti di denti non avevano una struttura primitiva secondo quanto sostenuto dagli evoluzionisti. Che il Liaoningornis avesse le caratteristiche dell'uccello fu affermato sulla rivista Discover, in un articolo che si chiedeva "Da dove provengono gli uccelli? Questo fossile suggerisce che non vengono dal ceppo dei dinosauri". 108
Un altro fossile che contraddice la tesi evoluzionista sull'Archaeopteryx è l'Eoalulavis. La struttura alare dell'Eoalulavis, che si disse fosse più giovane di 25-30 milioni di anni rispetto all'Archaeopteryx, è stata osservata nei moderni uccelli che volano lentamente.109 Ciò provò che 120 milioni di anni fa c’erano uccelli indistinguibili per molti aspetti da quelli che volano oggi nei cieli.
Questi fatti ancora una volta indicano che né l'Archaeopteryx né gli altri uccelli antichi simili rappresentarono forme di transizione. I fossili non indicano che le differenti specie di uccelli si siano evolute le une dalle altre. Al contrario, i reperti fossili provano che gli uccelli attuali e alcuni arcaici come l’Archaeopteryx sono vissuti nello stesso periodo. È vero che alcune specie di uccelli come l'Archaeopteryx e il Confuciusornis, si estinsero, ma il solo fatto che solo alcune delle specie preesistenti siano state in grado di sopravvivere sino ai giorni nostri non sostiene in se stesso la teoria dell’evoluzione.
Archaeoraptor: L’inganno dell’uccello-dinosauro
Incapaci di trovare nell’Archaeopteryxquello che cercavano, i sostenitori della teoria dell’evoluzione aggrapparono le loro speranze sugli altri fossili degli anni Novanta e su una serie di cosiddetti fossili di “uccelli-dinosauri” citati nei media mondiali. Si scoprì subito, però, che queste affermazioni erano semplicemente errate interpretazioni o peggio falsificazioni.
Il primo riferimento all’uccello-dinosauro fu la storia del fossile di “dinosauro pennuto dissotterrato in Cina” nel 1996, con gran clamore nei media. Fu trovato un fossile rettile chiamato Sinosauropteryx, ma alcuni paleontologi evoluzionisti che lo esaminarono dissero che aveva penne di uccello, a differenza dei rettili conosciuti. Esami condotti un anno dopo, però, dimostrarono che il fossile in realtà non aveva alcuna struttura simile alle penne di un uccello. Un articolo di Science dal titolo "Plucking the Feathered Dinosaur" affermava che le strutture dette “penne” dai paleontologi evoluzionisti non avevano assolutamente niente a che fare con le penne:

Una rappresentazione di Confuciusornis
Esattamente un anno fa, i paleontologi fecero molto rumore attorno alle foto di un cosiddetto “dinosauro pennuto” che giravano nelle sale del convegno annuale della Società di Paleontologia dei Vertebrati. L’esemplare di Sinosauropteryx proveniente dalla Formazione di Yixian in Cina era sulla copertina del New York Times e fu visto da alcuni come conferma delle origini degli uccelli dai dinosauri. Ma nel convegno sulla paleontologia dei vertebrati di quest’anno a Chicago, il mese scorso, il verdetto fu un po’ diverso. Le strutture non sono penne moderne, afferma la mezza dozzina di paleontologi occidentali che hanno visto l’esemplare. Il paleontologo dell’Università del Kansas, Larry Martin Lawrence, pensa che le strutture siano fibre di collagene sfilacciate sotto la pelle – e quindi, non hanno niente a che fare con gli uccelli.110
Un caso ancora più sensazionale di montatura sui dino-uccelli venne fuori nel 1999. Nel numero di novembre 1999, il National Geographic pubblicò un articolo su un esemplare fossile dissotterrato in Cina che si sosteneva avesse caratteristiche sia degli uccelli che dei dinosauri. L’autore dell'articolo del National Geographic, Christopher P. Sloan, si spinse fino ad affermare “ora possiamo dire che gli uccelli sono teropodi con la stessa sicurezza con cui affermiamo che gli esseri umani sono mammiferi”. A questa specie, che si diceva fosse vissuta 125 milioni di anni fa, fu immediatamente dato il nome scientifico di Archaeoraptor liaoningensis.111
Il fossile, però, era un falso ed era stato abilmente costruito assemblando cinque esemplari distinti. Un gruppo di ricercatori, tra cui anche tre paleontologi, dimostrò l'imbroglio un anno dopo con l'aiuto di una tomografia a raggi X. Il dino-uccello era in realtà il prodotto di un evoluzionista cinese. Gli improvvisati artigiani cinesi avevano formato il dino-uccello usando colla e cemento su 88 ossa e pietre. La ricerca suggerisce che l’Archaeoraptor sia stato costruito usando la parte anteriore dello scheletro di un antico uccello e che corpo e coda comprendessero ossa di quattro diversi esemplari.
La cosa interessante è che il National Geographic non solo pubblicò un articolo di alto profilo su un così evidente inganno, ma lo usò come base per affermare che gli scenari dell’ “evoluzione degli uccelli” erano stati verificati, senza esprimere alcun dubbio o cautela. Il dottor Storrs Olson, del famoso Museo di storia naturale dello Smithsonian Institute, negli Stati Uniti, disse in seguito di aver avvertito in anticipo che il fossile era un falso, ma che la redazione della rivista lo aveva totalmente ignorato. Secondo Olson, il “National Geographic ha raggiunto il livello più basso impegnandosi in un giornalismo da tabloid sensazionalistico e senza fondamenta”112.
In una lettera che scrisse a Peter Raven del National Geographic, Olson descrive in modo molto dettagliato la vera storia della montatura del “dinosauro pennuto”, fin dal lancio con un precedente articolo del National Geographic pubblicato nel 1998:
Prima della pubblicazione dell’articolo del National Geographic del luglio 1998, “I dinosauri mettono le ali”, Lou Mazzatenta fotografo dell’articolo di Sloan, mi invitò alla National Geographic Society per esaminare le sue fotografie dei fossili cinesi e per commentare il taglio che si stava dando alla storia. All’epoca cercai di far sapere che esistevano punti di vista, solidamente fondati, alternativi a quello che il National Geographic intendeva presentare ma, alla fine, mi fu chiaro che il National Geographic non era interessato ad altro se non al dogma prevalente secondo cui gli uccelli si evolvettero dai dinosauri.

Il Confuciusornis, che viveva alla stessa epoca dell‘Archaeopteryx, ha molte somiglianze con gli uccelli odierni.
L’articolo di Sloan porta il pregiudizio a un livello completamente nuovo e consiste in gran parte di informazioni non verificabili e non documentate che “creano” le notizie piuttosto che riportarle. La sua sfrontata affermazione secondo cui “ora possiamo dire che gli uccelli sono teropodi con la stessa sicurezza con cui affermiamo che gli esseri umani sono mammiferi” non viene nemmeno suggerita come qualcosa che riflette le opinioni di un particolare scienziato o gruppo di scienziati e quindi appare come poco più che propaganda editoriale. Questa melodrammatica affermazione era stata già confutata da studi recenti di embriologia e morfologia comparativa che, naturalmente, non sono mai citati.
Quello che è ancora più importante, però, è che non è stato in realtà dimostrato che qualcuna delle strutture illustrate nell’articolo di Sloan, che si afferma essere penne, lo sia veramente. Dire che lo sono è poco più che un augurio presentato come fatto. L’affermazione riportata a pagina 103, secondo cui "strutture cave, pilifere caratterizzano le proto-penne”, non ha senso considerando che le proto-penne esistono solo come costrutto teorico cosicché la struttura interna di una di esse è ancora più ipotetica.
La montatura sui dinosauri pennuti nella mostra attualmente allestita presso la National Geographic Society è ancora peggiore; vi leggiamo, infatti, la spudorata affermazione che ci sono forti prove che una grande varietà di dinosauri carnivori avesse penne. Un modello dell’indiscusso dinosauro Deinonychus e illustrazioni di cuccioli di tirannosauro sono messi in mostra ricoperti di penne, ma essi sono tutti semplicemente immaginari e non hanno alcun posto al di fuori della fantascienza.
- Distinti saluti
- Storrs L. Olson
- Curatore della sezione uccelli
- Museo naturale di storia naturale
- Smithsonian Institution113
Questo caso rivelatore dimostra due fatti importanti. Primo, che ci sono persone che non hanno remore a ricorrere a falsificazioni nello sforzo di trovare prove per la teoria dell’evoluzione. Secondo, che riviste scientifiche popolari e di ottima reputazione, che hanno assunto la missione di imporre la teoria dell’evoluzione alla gente, sono perfettamente disposte a trascurare qualsiasi fatto che possa essere sconveniente o che abbia interpretazioni alternative. Cioè sono diventate poco più che strumenti di propaganda per diffondere la teoria dell’evoluzione. Non hanno un punto di vista scientifico, ma dogmatico, e compromettono consapevolmente la scienza per difendere la teoria dell’evoluzione cui sono tanto fortemente devote.
Un grande scoop del National Geographic il “dino-uccello”. Fu subito chiaro che lo Archaeoraptor era una burla
Un altro aspetto importante della faccenda è che non ci sono prove per la tesi secondo cui gli uccelli si sarebbero evoluti dai dinosauri. In mancanza di prove, o si producono prove false o si interpretano male quellevere. In verità non c’è alcuna prova che gli uccelli si siano evoluti da altre specie viventi. Al contrario, tutte le scoperte dimostrano che gli uccelli sono comparsi sulla terra già in pieno possesso delle loro distinte strutture corporee.
L’origine degli insetti
Nel discutere l’origine degli uccelli abbiamo citato la teoria cursoria proposta dai biologi evoluzionisti. Come è stato chiarito, la questione del modo in cui i rettili svilupparono le ali comporta speculazioni su “rettili che cercavano di catturare insetti con le zampe anteriori”. Secondo questa teoria, le zampe anteriori di questi rettili si trasformarono lentamente in ali, nel corso del tempo, mentre essi cacciavano gli insetti.
Abbiamo già sottolineato che questa teoria non si basa su alcuna scoperta scientifica di sorta. Ma c’è un altro aspetto interessante che non abbiamo ancora toccato. Le mosche possono già volare. Come acquistarono le ali? È, parlando in genere, qual è l’origine degli insetti di cui le mosche sono solo una classe?
Nella classificazione degli esseri viventi gli insetti costituiscono un sub-phylum, gli insecta del phylum degli Arthropoda. l più antichi insetti fossili appartengono al periodo Devoniano (da 410 a 360 milioni di anni fa). Nel periodo Pennsylvaniano che seguì (da 325 a 286 milioni di anni fa), comparve un gran numero di specie di insetti diversi. Per esempio, le blatte comparvero all’improvviso e con le stesse strutture che hanno oggi. Betty Faber, del Museo Americano di Storia Naturale, dice che i fossili di blatte di 350 milioni di anni fa sono identici alle blatte di oggi.114
Creature come ragni, zecche e millepiedi non sono insetti, ma appartengono piuttosto ad altri sub-phyla degli artropodi. Importanti scoperte fossili di queste creature furono riportate nel 1983 nel corso del convegno annuale della American Association for the Advancement of Science. La cosa interessante riguardo a questi fossili di ragni, zecche e centopiedi vecchi di 350 milioni di anni è che non sono diversi dagli esemplari che vivono oggi. Uno degli scienziati che esaminò i fossili notò “sembra come se fossero morti ieri”.115
Insetti alati comparvero all’improvviso nei reperti fossili e da quel momento possiedono la stessa struttura perfetta di oggi. Il fossile di libellula di 320 milioni di anni che precede è l’esemplare più antico che si conosca e non è diverso dalle libellule che vivono oggi. Non c’è stata alcuna “evoluzione”.


Questo millepiedi Acantherpestes major, trovato nello stato del Kansas, negli Stati Uniti, ha circa 300 milioni di anni e non è diverso dai millepiedi odierni,.
Una mosca fossile di 145 milioni di anni. Il fossile, trovato a Liaoning, in Cina, è identico alle specie di mosche che vivono oggi.
Anche insetti alati emersero all’improvviso nei reperti fossili e con tutte le loro caratteristiche peculiari. Per esempio, è stato trovato un gran numero di fossili di libellule del periodo Pennsylvaniano. E tutte queste libellule hanno esattamente le stesse strutture dei loro omologhi di oggi.

Non c’è alcuna differenza tra questa blatta fossile di 320 milioni di anni e gli esemplari che vivono oggi.
Un punto interessante è che libellule e mosche comparvero all’improvviso insieme a insetti privi di ali. Questo confuta la teoria secondo cui insetti privi di ali le avrebbero sviluppate diventando gradualmente insetti volanti. In uno dei loro articoli riportati nel libro Biomechanics in Evolution, Robin Wootton e Charles P. Ellington dicono quanto segue sull’argomento:
Quando compaiono per la prima volta i fossili di insetti, nel Carbonifero medio e superiore, essi sono diversi e per la maggior parte completamente forniti di ali. Ci sono alcune forme primitive prive di ali ma non si conosce alcun intermedio convincente.116
Una caratteristica importante delle mosche, che comparvero all’improvviso nei reperti fossili, è la loro stupefacente tecnica di volo. Mentre un essere umano non può aprire e chiudere le braccia nemmeno dieci volte al secondo, una mosca, in media, sbatte le ali 500 volte nello stesso lasso di tempo. Inoltre, muove entrambe le ali contemporaneamente. La minima differenza tra la vibrazione delle ali farebbe perdere l’equilibrio alla mosca ma ciò non avviene mai.
In un articolo intitolato "The Mechanical Design of Fly Wings" Wootton osserva inoltre:
Meglio comprendiamo il funzionamento delle ali degli insetti, più ingegnosa e bella appare la loro struttura. Le strutture sono tradizionalmente disegnate in modo tale da deformarsi il meno possibile; i meccanismi sono progettati per muovere le parti che li compongono in maniera prevedibile. Le ali degli insetti combinano queste due caratteristiche, utilizzando componenti dotate di una vasta gamma di proprietà elastiche, elegantemente assemblate per permettere deformazioni in grado di rispondere alle forze che agiscono su di esse facendo uso dell'aria nel modo migliore. Tuttavia hanno pochi, o nessun parallelo tecnologico. 117
Naturalmente, l’improvviso apparire di esseri viventi con una struttura così perfetta non può essere spiegato su base evoluzionista. Ecco perché Pierre-Paul Grassé dice: “siamo al buio per quanto riguarda l’origine degli insetti.” 118 L’origine degli insetti dimostra il fatto che tutti gli esseri viventi furono creati da Dio.
Una mosca fossile, intrappolata nell’ambra 35 milioni di anni fa. Questo fossile, trovato sulla costa baltica, ancora una volta non è diverso da quelle che vivono ai giorni nostri.
L’origine dei mammiferi
Come affermato in precedenza, la teoria dell’evoluzione propone alcune creature immaginarie che sarebbero provenute dal mare e diventate rettili e che gli uccelli si sarebbero evoluti dai rettili. Secondo lo stesso scenario, i rettili sarebbero gli antenati non solo degli uccelli ma anche dei mammiferi. Ci sono però grandi differenze tra queste due classi. I mammiferi sono animali a sangue caldo (questo significa che generano il proprio calore e lo mantengono a livello costante), generano prole viva, allattano i loro piccoli e hanno il corpo ricoperto di pelliccia o peli. I rettili, d’altro canto, sono a sangue freddo (cioè non sono in grado di generare calore e la loro temperatura corporea cambia a seconda della temperatura esterna), depongono uova, non allattano i piccoli e hanno corpi ricoperti di scaglie.
Date tutte queste differenze, in che modo un rettile inizia a regolare la temperatura corporea e sviluppa un sistema di traspirazione che gli consente di mantenere la temperatura corporea? È possibile che abbia sostituito le scaglie con pelliccia e peli e iniziato a produrre latte? Perché la teoria dell’evoluzione spieghi l’origine dei mammiferi deve prima fornire risposte scientifiche a queste domande.
Tuttavia, quando guardiamo le fonti evoluzioniste, o troviamo scenari completamente immaginari e non scientifici o un profondo silenzio. Uno di questi scenari è quello che segue:
alcuni rettili delle regioni più fredde cominciarono a sviluppare un metodo per tenere caldi i loro corpi. La loro produzione di calore aumentava quando faceva freddo e la loro perdita di calore diminuì quando le scaglie diventarono più piccole e più appuntite e si evolvettero in pelliccia. Anche la sudorazione fu un adattamento per regolare la temperatura corporea, un modo per raffreddare il corpo, quando serviva, attraverso l’evaporazione dell’acqua. Ma incidentalmente, i cuccioli di questi rettili cominciarono a succhiare il sudore delle madri come nutrimento. Certe ghiandole sudorifere cominciarono a produrre una secrezione sempre più ricca che, alla fine, divenne latte. Così i cuccioli di questi primi mammiferi ebbero un inizio di vita migliore.119
Il suddetto scenario non è altro che un parto dell’immaginazione. Un tale fantastico scenario non solo non è sostenuto da prove, ma è chiaramente impossibile. È del tutto irrazionale affermare che una creatura vivente possa produrre un nutriente così complesso come il latte leccando il sudore della madre.
Il motivo per cui vengono presentati tali scenari è che ci sono enormi differenze tra rettili e mammiferi. Un esempio delle barriere strutturali tra rettili e mammiferi è la loro struttura mascellare. La mandibola dei mammiferi consiste in un osso mascellare che contiene i denti. Nei rettili ci sono tre piccole ossa su entrambi i lati della mandibola. Un’altra differenza fondamentale è che tutti i mammiferi hanno tre ossa nell’orecchio medio (martello, incudine e staffa). I rettili hanno un solo osso nell’orecchio medio. Gli evoluzionisti affermano che la mascella e l’orecchio medio dei rettili si evolvettero diventando la mascella e l’orecchio dei mammiferi. La domanda sul modo in cui un orecchio con un solo osso si sia potuto trasformare in uno con tre ossa e in che modo il senso dell’udito abbia potuto conservare nel frattempo il suo funzionamento non potrà mai trovare una risposta. Non sorprende che non sia stato trovato un solo fossile che colleghi i rettili e i mammiferi. Ecco perché Roger Lewin fu costretto a dire “la transizione al primo mammifero…è ancora un enigma”120.
George Gaylord Simpson, una delle più importanti autorità evoluzioniste e uno dei fondatori della teoria neodarwinista, fa il seguente commento a questa difficoltà imbarazzante per gli evoluzionisti:
l’evento che rende più perplessi nella storia della vita sulla terra è il cambiamento dal Mesozoico, l’era dei rettili, all’era dei mammiferi. È come se il sipario si fosse abbassato all’improvviso sulla fase in cui tutti i ruoli principali erano assunti da rettili, soprattutto dinosauri, in gran numero e stupefacente varietà, e si fosse sollevato di nuovo immediatamente per rivelare lo stesso scenario, ma con un nuovo cast, un cast in cui i dinosauri non recitano affatto e tutti gli altri rettili sono semplici comparse, mentre tutti i ruoli principali sono ricoperti da mammiferi di tipi a cui si faceva a stento cenno negli atti precedenti.121
Inoltre, quando i mammiferi fecero improvvisamente la loro comparsa, erano già diversi gli uni dagli altri. Questi animali diversi come pipistrelli, cavalli, topi e balene sono tutti mammiferi e comparvero tutti durante la stessa era geologica. Stabilire una relazione evolutiva tra di loro è impossibile anche con la più sfrenata immaginazione. Lo zoologo evoluzionista R. Eric Lombard sottolinea ciò in un articolo che apparve sull’importante rivista Evolution:
quelli che cercano specifiche informazioni utili a costruire la filogenesi dei taxa dei mammiferi saranno delusi.122
Non c’è alcuna differenza tra i mammiferi fossili di decine di milioni di anni fa presenti nei musei di storia naturale e quelli che vivono oggi. Inoltre questi fossili comparvero all’improvviso senza alcun collegamento a specie precedenti.
In breve, l’origine dei mammiferi, come quella di altri gruppi, non si conforma in alcun modo alla teoria dell’evoluzione. George Gaylord Simpson lo ammise molti anni fa:
questo è vero per tutti i trentadue ordini di mammiferi... I membri più antichi e primitivi che si conoscono di ogni ordine [di mammiferi] hanno già i caratteri di base dell’ordine e in nessun caso si conosce una sequenza approssimativamente continua da un ordine all’altro. Nella maggior parte dei casi, l'interruzione è così netta e la differenza tanto grande che l’origine dell’ordine è speculativa e molto discussa. Questa regolare assenza di forme di transizione non si limita ai mammiferi, ma è un fenomeno quasi universale, come è stato a lungo notato dai paleontologi. È vero per quasi tutte le classi di animali, sia vertebrati che invertebrati…è vero per le classi e per tutti i maggiori phyla animali ed è evidentemente vero anche per categorie analoghe di piante. 123
Il mito dell’evoluzione del cavallo
Un argomento importante nell’origine dei mammiferi è il mito dell’ “evoluzione del cavallo”, anch’esso un argomento a cui le pubblicazioni evoluzioniste hanno dedicato un notevole spazio per lungo tempo. Questo è un mito, perché si basa sull’immaginazione piuttosto che su scoperte scientifiche.
Fino a tempi recenti, un'immaginaria sequenza che si supponeva mostrasse l'evoluzione del cavallo, era presentata come la testimonianza fossile principale della teoria dell’evoluzione. Oggi, tuttavia, molti evoluzionisti ammettono francamente che lo scenario dell’evoluzione del cavallo ha fallito. Nel 1980, si tenne un simposio presso il Field Museum of Natural History di Chicago con la presenza di 150 evoluzionisti, per discutere i problemi della teoria evoluzionista gradualistica. Rivolgendosi ai convenuti, l’evoluzionista Boyce Rensberger notò che lo scenario dell’evoluzione del cavallo non ha fondamento nei reperti fossili e che non è stato osservato alcun processo evolutivo che spieghi la graduale evoluzione dei cavalli.
L’esempio comunemente raccontato dell'evoluzione del cavallo, che suggerisce una graduale sequenza di cambiamenti da una creatura con quattro dita grande quanto una volpe che viveva circa 50 milioni di anni fa, fino all'animale di oggi, molto più grande e con un solo dito, è stato riconosciuto già da molto tempo come errato. Invece di cambiamenti graduali, i fossili di ciascuna specie intermedia appaiono completamente distinti, persistono immutati e quindi si estinguono. Le forme di transizione sono sconosciute.124
La mostra L'evoluzione del Cavallo nel Museo di Storia Naturale di Londra. Questo e altri diagrammi dell’ “evoluzione del cavallo” mostrano specie indipendenti che vissero in epoche e luoghi diversi, allineati uno dopo l’altro in una presentazione molto soggettiva. In realtà non ci sono scoperte scientifiche riguardanti l’evoluzione del cavallo.
Trattando in modo particolarmente onesto questo importante dilemma nello scenario dell’evoluzione del cavallo, Rensberger mise in agenda la difficoltà della forma di transizione come la maggiore tra tutte le difficoltà.
Il dottor Niles Eldredge ha dichiarato quanto segue circa i diagrammi dell’“evoluzione del cavallo”:
C’è stata una terribile quantità di storie, alcune più fantasiose di altre, circa la reale natura [della vita].L'esempio più famoso, ancora in mostra al piano inferiore, è l’esposizione sull'evoluzione del cavallo, preparata forse cinquant'anni fa. È stata presentata come l'esatta verità, testo dopo testo. Ora io penso che ciò sia deplorevole, in particolare quando coloro che propongono questo tipo di storie possono essere consapevoli della natura speculativa di alcune di esse.125
Qual è allora lo scenario dell'evoluzione del cavallo? Questo scenario fu formulato attraverso diagrammi ingannevoli ipotizzati sulla base della sistemazione sequenziale di fossili di specie distinte che, secondo la grande forza d'immaginazione degli evoluzionisti, sarebbero vissute in periodi molto diversi in India, Sudafrica, Nord America ed Europa. Più di venti diagrammi sull'evoluzione del cavallo, che comunque sono completamente diversi l’uno dall’altro, sono stati proposti da vari ricercatori. Quindi è ovvio che gli evoluzionisti non hanno raggiunto un accordo su questi alberi genealogici. Il solo punto in comune in questi scenari è la credenza secondo cui una creatura delle dimensioni di un cane, detta Eohippus (Hyracotherium), che viveva nel periodo Eocene 55 milioni di anni fa, fosse il progenitore del cavallo. Il fatto è, però, che l‘Eohippus, che si estinse milioni di anni fa, è quasi identico all’irace, un piccolo animale simile al coniglio che ancora vive in Africa, e non ha niente a che vedere con il cavallo.126
L’inconsistenza della teoria dell'evoluzione del cavallo divenne sempre più evidente man mano che si raccoglievano reperti fossili. Fossili di specie di cavalli odierni (Equus nevadensis e Equus occidentalis) sono stati scoperti negli stessi strati dell‘Eohippus.127 Questa è una indicazione del fatto che il cavallo di oggi e il cosiddetto antenato vivevano allo stesso tempo.
Lo scrittore scientifico evoluzionista Gordon R. Taylor spiega questa verità poco conosciuta nel suo libro The Great Evolution Mistery:
Ma forse la più grave debolezza del darwinismo è la mancanza di paleontologi in grado di trovare filogenesi convincenti o sequenze di organismi capaci di dimostrare i maggiori cambiamenti evolutivi... Il cavallo è spesso citato come l'unico esempio compiuto. Ma il fatto è che la linea dall'Eohippus all'Equus è molto irregolare. È addotta per mostrare un continuo incremento di dimensioni, ma la realtà è che alcune varianti erano più piccole dell'Eohippus, non più grandi. Esemplari provenienti da fonti differenti possono essere riuniti in una sequenza all'apparenza convincente, ma non vi è prova sufficiente a confermare che essi fossero disposti secondo questo ordine temporale.128
Tutti questi fatti sono prove evidenti che i diagrammi sull'evoluzione del cavallo, presentati come una delle più solide dimostrazioni della teoria dell’evoluzione, non sono altro che favole fantastiche e poco plausibili. Come altre specie, anche i cavalli giunsero all’esistenza senza antenati in senso evolutivo.
L’origine dei pipistrelli

Il sistema di sonar dei pipistrelli è più sensibile ed efficiente di qualsiasi sistema di sonar tecnologico costruito finora.
Una delle creature più interessanti nella classe dei mammiferi è senza dubbio il mammifero volante, il pipistrello.
In cima alla lista delle caratteristiche dei pipistrelli c’è il complesso sistema di sonar che possiedono. Grazie a esso, i pipistrelli possono volare nel buio fitto, incapaci di vedere alcunché ma eseguendo le manovre più complicate. Possono perfino percepire e catturare un bruco sul pavimento di una stanza buia.
I sonar dei pipistrelli funzionano come segue. L’animale emette un flusso continuo di segnali sonici ad alta frequenza, analizza gli echi che ne provengono e, come risultato, forma un’immagine dettagliata di quanto lo circonda. Per di più, riesce a fare tutto ciò a una velocità incredibile, in modo continuo e senza errori, mentre vola in aria.
Le ricerche sul sistema di sonar dei pipistrelli hanno prodotto risultati ancora più sorprendenti. La gamma di frequenze che l’animale può percepire è molto stretta, in altre parole può udire solo suoni di certe frequenze, e qui abbiamo un punto molto importante. Poiché i suoni che colpiscono un corpo in movimento cambiano frequenza (il ben noto “effetto Doppler”) appena un pipistrello manda segnali ad una mosca, ad esempio, che si allontana da lui volando, le onde sonore riflesse dalla mosca dovrebbero essere a una frequenza diversa che il pipistrello non è in grado di percepire. Per questo motivo, il pipistrello dovrebbe avere grandi difficoltà a percepire corpi in movimento.
Ma non è così. Il pipistrello continua a catturare ogni tipo di piccola creatura che si muove velocemente senza alcuna difficoltà. Il motivo è che il pipistrello regola la frequenza delle onde sonore che invia verso i corpi in movimento nell’ambiente come se sapesse tutto sull’effetto Doppler. Per esempio, emette il segnale di massima frequenza verso una mosca che si allontana da lui in modo che, quando il segnale torna, la frequenza non è scesa al di sotto della soglia dell'udito dell'animale.
In che modo avviene questa regolazione?
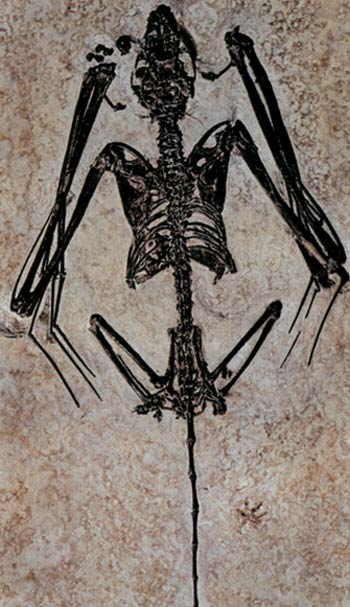
Il pipistrello fossile più antico che si conosca, trovato nel Wyoming negli Stati Uniti. Vecchio di 50 milioni di anni, non c’è differenza tra questo fossile e i pipistrelli che vivono oggi.
Ci sono due gruppi di neuroni (cellule nervose) nel cervello del pipistrello che controllano il sistema sonar. Uno di questi percepisce gli ultrasuoni echeggiati e l’altro dà istruzioni ai muscoli per produrre chiamate di ecolocazione. Queste regioni del cervello operano in coppia, in modo che quando la frequenza dell’eco cambia, la prima regione lo percepisce e avverte la seconda, rendendo possibile modificare la frequenza del suono emesso secondo quella dell’eco. Di conseguenza il passo dell’ultrasuono del pipistrello cambia secondo quello che lo circonda e il sistema sonar nel complesso viene usato nella maniera più efficiente.
È impossibile non vedere il colpo mortale che il sistema sonar del pipistrello infligge ala teoria dell’evoluzione graduale attraverso mutazioni casuali. È una struttura estremamente complessa e non può in alcun modo essere spiegata con mutazioni casuali. Perché il sistema funzioni, tutti i componenti devono operare perfettamente assieme come un tutto integrato. È assurdo credere che un sistema così altamente integrato possa essere spiegato dal caso, al contrario dimostra in realtà che il pipistrello è stato creato perfettamente.
In realtà anche i reperti fossili confermano che i pipistrelli comparvero immediatamente e con le complesse strutture odierne. Nel loro libro Bats: A Natural History, i paleontologi evoluzionisti John E. Hill e James D. Smith rivelano questo fatto facendo la seguente ammissione:
i reperti fossili di pipistrelli arrivano fino al primo Eocene e sono documentati nei cinque continenti…[Tutti] i pipistrelli fossili, anche i più antichi, sono chiaramente completamente sviluppati e quindi gettano poca luce sulla transizione dai loro antenati terrestri.129
E il paleontologo evoluzionista L. R. Godfrey dice questo sullo stesso argomento:
ci sono alcuni pipistrelli fossili del primo Terziario, notevolmente ben conservati, come gli Icaronycteris ma essi non ci dicono alcunché sull’evoluzione del volo nei pipistrelli perché erano pipistrelli che volavano perfettamente bene.130
Lo scienziato evoluzionista Jeff Hecht confessa lo stesso problema in un articolo comparso su New Scientist del 1998:
[le] origini dei pipistrelli sono un enigma. Anche i più antichi pipistrelli fossili, di circa 50 milioni di anni fa, hanno ali che somigliano molto da vicino a quelle dei pipistrelli moderni.131
In breve, il complesso sistema corporeo dei pipistrelli non può essere emerso attraverso l’evoluzione e i reperti fossili dimostrano che ciò non avvenne. Al contrario, i primi pipistrelli comparsi nel mondo sono esattamente gli stessi di oggi. I pipistrelli sono sempre esistiti come pipistrelli.
L’origine dei mammiferi marini
Balene e delfini appartengono all’ordine dei mammiferi marini noti come Cetacei. Queste creature sono classificate come mammiferi perché, come i mammiferi che vivono sulla terra, danno alla luce una prole viva, la nutrono, hanno polmoni per respirare e regolano la temperatura corporea. Per gli evoluzionisti, l’origine dei mammiferi marini è stata una delle più difficili da spiegare. In molte fonti evoluzioniste si afferma che gli antenati dei cetacei avrebbero lasciato la terra e si sarebbero evoluti in mammiferi marini nel corso di un lungo periodo di tempo. Di conseguenza, i mammiferi marini avrebbero seguito un percorso contrario alla transizione dall’acqua alla terra e avrebbero subito un secondo processo evolutivo, tornando nell’acqua. Questa teoria manca di prove paleontologiche ed è contraddittoria in sé. Quindi gli evoluzionisti sono costretti da molto tempo a tacere su questo tema.
Ma una montatura evoluzionista sulle origini dei mammiferi marini, venuta fuori negli anni Novanta, affermò di basarsi su due nuovi fossili, Pakicetus e Ambulocetus, scoperti negli anni Ottanta. Questi mammiferi estinti, evidentemente quadrupedi e terrestri, furono presentati come presunti antenati delle balene e quindi molte fonti evoluzioniste non esitarono a chiamarli “balene ambulanti”. (in realtà il nome completo, Ambulocetus natans, significa “balena che cammina a nuota”). Popolari mezzi di indottrinamento evoluzionista, inoltre, diffusero la storia. Il National Geographic, nel suo numero del novembre 2001, descrisse il completo scenario evoluzionista sulla “evoluzione delle belene”, o scenario, però, si basava sul pregiudizio evoluzionista, non su prove scientifiche.
Il mito della balena ambulante
Resti fossili del mammifero estinto, Pakicetus inachus, per chiamarlo col suo vero nome, entrarono in scena per la prima volta nel 1983. P. D. Gingerich e i suoi assistenti, che avevano trovato il fossile, non ebbero esitazioni nell’affermare immediatamente che era un “balena primitiva”, anche se in realtà avevano trovato solo un cranio.
Il fossile, tuttavia, non ha assolutamente alcun collegamento con la balena. Lo scheletro si rivelò essere una struttura a quattro zampe simile a quella del lupo comune. Fu trovato in una regione piena di minerali ferrosi e contenente fossili di creature terrestri come serpenti, tartarughe e coccodrilli. In altre parole, faceva parte di uno strato di terra, non di uno acquatico.
Allora in che modo un quadrupede abitante sulla terra veniva annunciato come una “balena primitiva”? Solo basandosi su alcuni dettagli dei denti e delle ossa dell’orecchio! Queste caratteristiche, però, non sono una prova su cui basare il collegamento tra il Pakicetus e la balena.
Anche gli evoluzionisti ammettono che la maggior parte della relazione teorica costruita sulla base delle somiglianze anatomiche tra animali è del tutto inaffidabile. Se l’ornitorinco, un mammifero dotato di becco, e l’anatra fossero entrambi estinti da molto tempo, allora senza dubbio gli evoluzionisti li definirebbero parenti stretti sulla base del becco simile. Ma poiché l’ornitorinco è un mammifero e l’anatra un uccello, la teoria dell'evoluzione non può stabilire alcun collegamento nemmeno tra di loro,
Il Pakicetus, che gli evoluzionisti dichiarano essere una “balena ambulante”, era una specie unica che ospitava diverse caratteristiche nel suo corpo. Infatti, Carroll, un’autorità nel campo della paleontologia dei vertebrati, descrive la famiglia dei mesonichidi, di cui il Pakicetusdovrebbe essere membro, visto “che presenta una strana combinazione di caratteri”132. Anche un eminente evoluzionista come Gould ammette che tali “creature a mosaico” non possono essere considerate forme evolutive intermedie.
Nel suo articolo "The Overselling of Whale Evolution", lo scrittore creazionista Ashby L. Camp rivela la totale mancanza di validità dell’affermazione secondo cui la classe dei Mesonichidi, che dovrebbe comprendere mammiferi terrestri come il Pakicetus, potrebbe essere stata antenata degli Archaeoceti, una specie estinta di balene, con queste parole:
Il motivo per cui gli evoluzionisti credono che i mesonichidi dettero origine agli archaeoceti, nonostante l’impossibilità di identificare alcuna specie nella linea attuale, è che mesonichidi e archaeoceti noti hanno alcune similitudini. Queste similitudini, però, non sono sufficienti a farne antenati, specialmente alla luce delle grosse differenze. La natura soggettiva di tali confronti è resa evidente dal fatto che tanti gruppi di mammiferi e persino di rettili sono stati indicati come antenati della balene.133
Ambulocetus natans: una falsa balena con artigli “palmati”
La seconda creatura fossile nello scenario delle origini delle balene, dopo il Pakicetus, è l‘Ambulocetusnatans. È in realtà una creatura terrestre che gli evoluzionisti hanno invece trasformato in una balena.
Il nome deriva dalle parole latine “ambulare”, “cetus” e “natans” e significa “una balena che cammina e nuota” È ovvio che l’animale camminava perché aveva quattro zampe come tutti i mammiferi terrestri e persino larghi artigli e piedi sulle zampe posteriori. A parte il pregiudizio evoluzionista, però, non c’è assolutamente alcuna base per affermare che nuotasse nell’acqua o che vivesse sia sulla terra che nell’acqua (come gli anfibi).

La ricostruzione del Pakicetus da parte del National
Geographic.
DISTORSIONI NELLE RICOSTRUZIONI DEL NATIONAL GEOGRAPHIC
I paleontologi credono che il Pakicetus era un mammifero quadrupede. La struttura dello scheletro a sinistra (in alto), pubblicata nella rivista Nature (vol. 412, 20 settembre 2001) lo dimostra chiaramente. Quindi la ricostruzione del Pakicetus (al centro) fatta da Carl Buell, che si basa su questa struttura, è realistica.
Il National Geographic, però, scelse di usare un‘immagine di un Pakicetus “nuotatore” (in basso) per descrivere l’animale come una “balena ambulante” e imporre tale immagine ai suoi lettori. Le incoerenze dell’immagine, destinate a far sembrare il Pakicetus “simile alla balena” sono ovvie. L’animale è stato ritratto in una posizione “di nuoto”. Le zampe posteriori sono mostrate allungate all’indietro ed è stata data un‘impressione di “pinne”.
Per vedere il confine tra la scienza e l'immaginazione su questo argomento, diamo uno sguardo alla ricostruzione che il National Geographic fa dell'Ambulocetus. Ecco come viene ritratto nella rivista:
Se si guarda attentamente, si possono facilmente vedere le due piccole manipolazioni che sono state utilizzate per trasformare l‘Ambulocetus in una balena.
• Le zampe posteriori dell’animale vengono mostrate non come piedi che gli avrebbero consentito di camminare, ma come pinne che lo avrebbero assistito nel nuoto. Tuttavia Carroll, che esaminò le ossa delle zampe dell’animale, dice che aveva la capacità di muoversi con sicurezza sulla terra.
• Per dare l’impressione che fossero adatte al nuoto, le zampe anteriori sono state disegnate palmate. È impossibile, però, trarre tali conclusioni da uno studio dei fossili di Ambulocetus. Nei reperti fossili è quasi impossibile trovare tessuti molli come questi. Quindi la ricostruzione basata su caratteristiche oltre a quelle dello scheletro sono sempre speculative. Questo offre agli evoluzionisti uno spazio vuoto molto ampio per speculazioni da usare come strumenti di propaganda.
Con lo stesso tocco evoluzionista applicato al disegno dell’Ambulocetus, è possibile far sì che qualunque animale somigli a un altro. Si può persino prendere lo scheletro di una scimmia, disegnare pinne sul dorso e far diventare le dita palmate e presentarlo come il “primo antenato delle balene”.
L’invalidità dell’inganno perpetrato sulla base del fossile di Ambulocetussi può vedere dai disegni che seguono, riportati nello stesso numero del National Geographic:
Nel pubblicare l’immagine dello scheletro dell’animale, il National Geographic dovette fare un passo indietro rispetto al ritocco che aveva apportato alla figura della ricostruzione che lo faceva somigliare di più a una balena. Come mostra chiaramente lo scheletro, le ossa del piede dell’animale erano strutturate per sostenerlo sulla terra. Non c’è alcun segno di immaginarie dita palmate.
L’inattendibilità del mito della balena ambulante
In verità non c’è alcuna prova che Pakicetuse Ambulocetus siano antenati della balena. Sono semplicemente descritti come “possibili antenati”, sulla base di alcune limitate somiglianze, da evoluzionisti smaniosi di trovare un antenato terrestre ai mammiferi marini, alla luce della loro teoria. Non c’è alcuna prova che colleghi queste creature ai mammiferi marini che appaiono nei reperti fossili in un tempo geologico molto simile.
Dopo Pakicetus e Ambulocetus, il progetto evoluzionista si sposta sui mammiferi marini e presenta specie estinte di balene come Procetus, Rodhocetus e Archaeocetea. Gli animali in questione erano mammiferi che vivevano nel mare e che ora sono estinti, un argomento su cui ritorneremo più avanti. Ci sono però notevoli differenze anatomiche tra questi e Pakicetus e Ambulocetus. Se osserviamo i fossili, è chiaro che non sono “forme di transizione” che li collegano l’uno all’altro.
• La spina dorsale del mammifero quadrupede termina nel bacino e nelle possenti zampe posteriori che si estendono da esso. Questa è la tipica anatomia dei mammiferi terrestri. Nelle balene, invece, la spina dorsale va fino alla coda e non c’è alcun osso pelvico. Infatti il Basilosaurus, che si ritiene sia vissuto circa 10 milioni di anni dopo l‘Ambulocetus, possedeva quest’ultima anatomia. In altre parole, è una tipica balena. Non c’è alcuna forma di transizione tra l‘Ambulocetus, un tipico mammifero terrestre e il Basilosaurus, una tipica balena.
• Sotto la colonna dorsale del Basilosauruse del capodoglio ci sono piccole ossa indipendenti da essa. Gli evoluzionisti sostengono che questi siano resti di zampe. Nel Basilosaurus però, queste ossa fungevano da guida per l’accoppiamento e nei capodogli “[agiscono] da ancoraggio per i muscoli dei genitali."135 Descrivere queste ossa come “organi vestigiali” non è altro che un pregiudizio darwinista.
In conclusione, nulla cambia il fatto che non c’era alcuna forma di transizione tra mammiferi terrestri e marini e che entrambi emersero con le loro caratteristiche particolari. Non c’è alcun collegamento evolutivo. Robert Carroll lo accetta, anche se a malincuore e nel linguaggio evoluzionista: “non è possibile identificare una sequenza di mesonichidi che porti direttamente alla balene”.136
Sebbene sia un evoluzionista, nemmeno il famoso esperto russo di balene G. A. Mchedlidze, sostiene la descrizione di Pakicetus, Ambulocetus natanse simili creature a quattro zampe come “possibili antenati della balena” e li descrive invece in un gruppo completamente isolato.137
Racconti evoluzionistici su orecchie e nasi
Qualsiasi scenario di un rapporto evolutivo tra mammiferi terrestri e marini deve spiegare le diverse strutture di orecchio e naso nei due gruppi. Per prima cosa consideriamo la struttura dell’orecchio. Come noi, i mammiferi di terra catturano i suoni che provengono dall’esterno nell’orecchio esterno, li amplificano con le ossa dell’orecchio medio e li trasformano in segnali nell’orecchio interno. I mammiferi marini non hanno orecchie. Odono i suoni attraverso recettori sensibili alle vibrazioni presenti nella mascella inferiore. Il punto cruciale è che è impossibile qualsiasi evoluzione per fasi tra un sistema uditivo perfetto verso uno completamente diverso. Le fasi di transizione non sarebbero vantaggiose. Un animale che perde lentamente la capacità di udire con le orecchie e non ha ancora sviluppato quella di udire attraverso la mascella, è in svantaggio.
Il problema del modo in cui potrebbe essere avvenuto tale “sviluppo” è un dilemma insolubile per gli evoluzionisti. I meccanismi che gli evoluzionisti presentano sono le mutazioni e queste non sono mai state viste aggiungere informazioni nuove e significative alle informazioni genetiche dell’animale. Non è ragionevole suggerire che il complesso sistema uditivo dei mammiferi marini possa essere emerso come risultati di mutazioni.
Infatti i fossili dimostrano che non è mai avvenuta alcuna evoluzione. Il sistema uditivo di Pakicetus e Ambulocetus è lo stesso dei mammiferi terrestri. Il Basilosaurus, che segue questi due mammiferi terrestri nell‘ipotetico “albero evolutivo”, d’altro canto possiede un tipico orecchio da balena. Era una creatura che percepiva i suoni attorno a sé, non attraverso un orecchio esterno ma attraverso le vibrazioni che raggiungevano la sua mascella. E non c’è alcuna “forma di transizione” tra l’orecchio del Basilosaurus e quello del Pakicetus e dell’Ambulocetus.
E’ simile il caso del racconto del “naso scorrevole”. Le fonti evoluzioniste dispongono in fila tre teschi di Pakicetus, di Rodhocetuse di una balena grigia dei giorni nostri e affermano che rappresentano un “processo evolutivo”. Tuttavia le tre strutture nasali dei fossili, specialmente quelle del Rodhocetuse della balena grigia, sono tanto diverse che è impossibile accettarle come forme di transizione nella stessa serie.
Inoltre il movimento delle narici verso la fronte richiederebbe una nuova strutturazione nell’anatomia degli animali in questione e credere che questo sia potuto avvenire come risultato di mutazioni casuali è una pura e semplice fantasia.
I racconti lamarckiani del National Geographic
Molti evoluzionisti sostengono un tipo di superstizione circa l’origine degli esseri viventi. Questa superstizione è la magica “forza naturale” che consentirebbe agli esseri viventi di acquisire organi, strutture biochimiche e caratteristiche anatomiche di cui hanno bisogno. Diamo uno sguardo ad alcuni interessanti brani dell’articolo "Evolution of Whales" comparso sul National Geographic:
…ho cercato di visualizzare alcune delle varietà di antenati della balena trovati qua e là… Man mano che le membra posteriori si rimpicciolivano o lo facevano anche le ossa dell’anca che le sostenevano...Il collo si accorciava trasformando l'estremità principale del corpo in una carena tubolare per solcare le acque con il minimo attrito, mentre le braccia assumevano la forma di timoni. Avendo poca necessità di orecchio esterno, alcune balene ricevevano i suoni prodotti in acqua direttamente attraverso la mascella inferiore e li trasmettevano all’orecchio interno attraverso speciali cuscinetti di grasso.138
A un‘attenta ispezione, in tutto questo resoconto la mentalità evoluzionista dice che gli esseri viventi sentono il bisognoi di cambiare, assecondando l’ambiente in trasformazione in cui vivono e questo bisogno è percepito come “meccanismo evolutivo”. Secondo questa logica, organi meno necessari scompaiono e organi necessari compaiono in armonia!
Chiunque abbia la più piccola conoscenza della biologia sa che i nostri bisogni non danno forma ai nostri organi in modo ereditario. Da quando fu confutata la teoria di Lamarck del passaggio delle caratteristiche acquisite alle generazioni successive, in altre parole da circa un secolo, questo è un fatto acquisito. Tuttavia, se si leggono le pubblicazioni evoluzioniste, esse sembrano pensare ancora secondo le linee di Lamarck. Se lo si fa notare, dicono: “no, noi non crediamo in Lamarck. Quello che diciamo è che le condizioni naturali pongono una pressione evolutiva sugli esseri viventi e, come conseguenza di ciò, le caratteristiche approvate vengono selezionate e, in questo modo, le specie si evolvono.” Qui però sta il punto critico: quella che gli evoluzionisti chiamano “pressione evolutiva” non può portare gli esseri viventi ad acquisire nuove caratteristiche secondo i loro bisogni. Questo perché i due cosiddetti meccanismi evolutivi che si suppone rispondano a questa pressione, la selezione naturale e la mutazione, non possono fornire nuovi organi agli animali.
• La selezione naturale può solo selezionare caratteristiche che già esistono, non crearne di nuove.
• Le mutazioni non possono aggiungere niente alle informazioni genetiche, possono solo distruggere quello che esiste. Non è stata mai osservata alcuna mutazione che aggiunga in modo inequivocabile informazioni significative al genoma (e che quindi formino un nuovo organo o una nuova struttura biochimica).
Se, alla luce di questo fatto, guardiamo ancora una volta al mito delle balene che si muovono goffamente, riportato nel National Geographic, vediamo che sono in realtà ancora impegnati in un lamarckismo primitivo. A un‘attenta analisi, lo scrittore Douglas H. Chadwick del National Geographic “visualizza” che le “membra posteriori si rimpicciolivano” in ciascuna balena della sequenza. In che modo una modifica morfologica potrebbe avvenire in una specie nel corso di generazioni, verso una direzione particolare? Perché questo avvenga, rappresentanti di quella specie, in ogni sequenza, avrebbero dovuto subire mutazioni per accorciare le zampe, tale mutazione non avrebbe dovuto causare alcun danno all’animale, questi mutanti avrebbero dovuto avere vantaggi rispetto agli animali normali, le successive generazioni, per una grande coincidenza, avrebbero dovuto subire la stessa mutazione nello stesso punto del gene, questo sarebbe dovuto passare immutato per molte generazioni e tutto ciò sarebbe dovuto avvenire per caso e del tutto senza errori.
Se gli scrittori del National Geographic credono ciò, allora credono anche a qualcuno che dice: “alla mia famiglia piace volare. Mio figlio ha subito una mutazione e sotto le sue braccia si sono sviluppate alcune strutture simili alle penne degli uccelli. Mio nipote subirà la stessa mutazione e le penne aumenteranno. Questo andrà avanti per generazioni e alla fine i miei discendenti avranno le ali e potranno volare". Entrambe le storie sono ugualmente ridicole.
Come detto all’inizio, gli evoluzionisti espongono la superstizione secondo cui i bisogni degli esseri viventi possono essere soddisfatti da una magica forza della natura. Il fatto di ascrivere consapevolezza alla natura, una credenza che si incontra nelle culture animiste, sta crescendo in modo interessante sotto i nostri occhi, nel ventunesimo secolo, sotto un‘aureola “scientifica”. Come il famoso biologo francese Paul-Pierre Grassé, un eminente critico del darwinismo, ha chiarito ancora una volta, però, "non c’è alcuna legge contro i sogni a occhi aperti ma la scienza non deve indulgere in essi”.139
Un altro scenario che gli evoluzionisti stanno cercando di imporre, senza molta discussione, riguarda la superficie corporea degli animali in questione. Come per altri mammiferi, si è d’accordo sul fatto che il Pakicetus e l’Ambulocetus, come mammiferi di terra, avevano il corpo coperto di pelliccia. E, nelle ricostruzioni, sono mostrati col corpo coperto di spessa pelliccia. Tuttavia, quando si passa ad animali successivi (veri mammiferi marini) tutta la pelliccia scompare. La spiegazione evoluzionista di ciò non è diversa dai fantastici scenari di tipo lamarckiano visti in precedenza.
La verità è che tutti gli animali in questione furono creati nella maniera più appropriata al loro ambiente. È irrazionale cercare di spiegarlo per mezzo di mutazioni o attraverso le facili storie lamarckiane. Come tutte le caratteristiche della vita, i sistemi perfetti di queste creature manifestano il fatto che furono creati da Dio.
Impasse dello scenario dell’evoluzione dei mammiferi marini.
Finora abbiamo esaminato quanto fallace sia lo scenario secondo cui i mammiferi marini si sarebbero evoluti da quelli di terra. Le prove scientifiche dimostrano che non esiste alcuna relazione tra i due mammiferi terrestri (Pakicetuse Ambulocetus), che gli evoluzionisti pongono all’inizio del racconto, e i mammiferi marini. E che dire del resto dello scenario?
La teoria dell’evoluzione è di nuovo in grande difficoltà. La teoria cerca di stabilire un collegamento fitogenico tra gli Archeoceti (balene arcaiche), mammiferi marini che si sa essere estinti, e balene e delfini viventi. La paleontologa evoluzionista Barbara J. Stahl, però, ammette che “la forma serpentina del corpo e i peculiari denti a sega rendono chiaro che questi Archeoceti non potevano essere stati antenati di alcuna delle balene dei nostri giorni.
La spiegazione evoluzionista delle origini dei mammiferi marini affronta un’enorme impasse sotto forma delle scoperte nel campo della biologia molecolare. Il classico scenario evoluzionista ipotizza che i due maggiori gruppi di balene, le balene dentate (Odontoceti) e le balene con fenoni (Misticeti), si siano evoluti da un antenato comune. Tuttavia Michel Milinkovitch dell’Università di Bruxelles ha opposto a questa opinione una nuova teoria. Egli ha sottolineato come questa ipotesi, che si basa su somiglianze anatomiche, si astata confutata dalle scoperte molecolari.
I rapporti evolutivi tra i maggiori gruppi di cetacei sono più problematici perché le analisi morfologiche e molecolari giungono a conclusioni molto diverse. In effetti, sulla base della convenzionale interpretazione dei dati morfologici e comportamentali, le balene dentate eco-localizzanti (circa 67 specie) e le balene con fenoni che filtrano il cibo (10 specie) sono considerate due gruppi morfologici distinti... D’altro canto l’analisi filogenetica del DNA e le sequenze degli aminoacidi contraddicono questa divisone tassonomica a lungo accettata. Un gruppo di balene dentate, i capodogli, sembra essere più strettamente imparentati alle balenottere - molto diverse dal punto di vista morfologico – che agli altri odontoceti.141
In breve, i mammiferi marini sfidano gli immaginari scenari evolutivi a cui sono costretti ad adattarsi.
Al contrario di quanto afferma la propaganda evoluzionista sulle origini dei mammiferi marini, non parliamo di un processo evolutivo supportato da prove scientifiche ma da prove costrette ad adattarsi al presupposto albero evolutivo, nonostante le molte contraddizioni tra di essi.
Quello che emerge, se si guarda obiettivamente alle prove, è che gruppi diversi di esseri viventi emersero indipendentemente l’uno dall’altro, nel passato. Questa è una rigorosa prova empirica del fatto che tutte queste creature furono create.
I mammiferi sono considerati come la forma di vita in cima alla cosiddetta scala evolutiva. Se è così, diventa difficile spiegare perchè questi animali si sarebbero spostati in un ambiente marino. Un’altra domanda è in che modo queste creature si sarebbero adattati all’ambiente marino anche meglio dei pesci, dal momento che animali come l’orca e il delfino, che sono mammiferi e quindi posseggono polmoni, si sono adattati all’ambiente in cui vivono anche meglio dei pesci che respirano nell’acqua.
È assolutamente ovvio che l’immaginaria evoluzione dei mammiferi marini non può essere spiegata in termini di mutazioni e selezione naturale. Un articolo pubblicato sulla rivista GEO fa riferimento alle origini della balena blu, un mammifero marino, e afferma così la disperata posizione del darwinismo sull’argomento:
Come per le balenottere azzurre, la struttura corporea e gli organi di altri mammiferi che vivono nel mare somigliano anch’essi a quelli dei pesci. Anche lo scheletro presenta somiglianze con quello dei pesci. Nelle balene, le membra posteriori che possono essere definite zampe, mostrano un’involuzione, non raggiungendo una crescita completa. Tuttavia non c’è la minima informazione circa modifiche nella forma di questi animali. Dobbiamo presumere che il ritorno al mare ebbe luogo non attraverso una lenta e lunga transizione come affermato dal darwinismo, ma avvenne in un salto istantaneo. Oggi i paleontologi non hanno sufficienti informazioni circa le specie di mammiferi da cui si evolvettero le balene.142
In effetti è molto difficile immaginare come un piccolo mammifero che viveva sulla terra asciutta si sia trasformato in una balena lunga 30 metri e pesante circa 60 tonnellate. Tutto quello che i darwinisti possono fare rispetto a ciò è produrre parti dell’immaginazione come nel seguente estratto di un articolo pubblicato dal National Geographic:
La superiorità di dimensioni della balena cominciò probabilmente sessanta milioni di anni fa, quando un peloso mammifero a quattro zampe si avventurò nell'acqua in cerca di cibo o rifugio. Col passare degli eoni, si ebbero dei lenti cambiamenti. Le zampe posteriori scomparvero, quelle anteriori diventarono pinne natatorie, il pelame lasciò il posto a uno spesso e liscio strato di grasso, le narici si spostarono alla sommità del capo, la coda si ampliò e galleggiando nell’acqua, il corpo divenne enorme. 143
Gli scenari della graduale evoluzione, descritti in precedenza, non soddisfano nessuno, nemmeno i loro stessi autori. Ma, in ogni caso, esaminiamo i dettagli di questo racconto fase per fase per vedere quanto irrealistico esso sia in realtà.
La peculiare struttura dei mammiferi marini
Per comprendere quanto sia impossibile lo scenario evoluzionista riguardo ai mammiferi marini, esaminiamo brevemente qualche altra caratteristica peculiare di questi animali. Quando si prendono in considerazione gli adattamenti che un mammifero terrestre deve subire per evolversi in un mammifero marino, anche la parola “impossibile” sembra inadeguata. Durante una tale transizione, anche se una sola delle fasi intermedie fosse mancata, la creatura non sarebbe potuta sopravvivere, il che avrebbe messo fine all’intero processo. Gli adattamenti che i mammiferi marini devono aver affrontato durante il passaggio verso l'acqua sono i seguenti:
LE GRANDI DIFFERENZE MORFOLOGICHE TRA ANIMALI CHE SI AFFERMA ESSERE DISCESI L’UNO DALL’ALTRO
Finora abbiamo visto specie diverse emergere sulla terra senza alcuna “forma intermedia” evolutiva tra loro. Esse appaiono nei reperti fossili con differenze talmente grandi che è impossibile stabilire un collegamento evolutivo fra di loro.
Se confrontiamo le loro strutture scheletriche, questo fatto si può vedere chiaramente ancora una volta. Animali che si diceva fossero parenti evolutivi differiscono enormemente. Ne esamineremo ora alcuni esempi. Tutti i disegni sono stati tratti da fonti evoluzioniste, da parte di esperti in vertebrati (come contestato anche da Michael Denton nel suo Evolution: A Theory in Crisis).

Il rettile marino
Mesosaurus ritenuto
un‘evoluzione
dell‘Hylonomus
Il rettile marino Ittiosauro
ritenuto una evoluzione dello
Hylonomus
L‘Hylonomus, il rettile marino più antico che si conosca.
Due diverse specie di rettili marini e l’animale terrestre che gli evoluzionisti affermano essere il loro antenato più prossimo. Notare le grandi differenze tra loro.
1- Ritenzione idrica: a differenza di altri animali marini, i mammiferi marini non possono usare l’acqua per soddisfare i loro bisogni idrici. Per sopravvivere hanno bisogno di acqua dolce. Anche se abbiamo informazioni limitate circa le risorse di acqua dolce dei mammiferi marini, si pensa che si alimentino di organismi che hanno una proporzione di sale relativamente bassa (circa un terzo di quella dell’acqua del mare). Quindi per i mammiferi marini la ritenzione dell’acqua nel corpo è cruciale. Ecco perché hanno un meccanismo di ritenzione idrica simile a quello dei cammelli. Come i cammelli, i mammiferi marini non sudano ma i loro reni sono perfettamente funzionanti producendo un‘urina altamente concentrata che consente all’animale di risparmiare acqua. In questo modo la perdita d’acqua si riduce al minimo.

Il Dimorphodon, uno dei rettili volanti più antichi che si conoscano, un tipico rappresentante di questo gruppo.
L‘Archaeopteryx, l’uccello
più antico che si conosca.
Il rettile di terra Euparkeria,
ritenuto da molte autorità
evoluzioniste l'antenato di
uccelli e rettili di terra.
Le plus vieil oiseau connu (archæoptéryx), un reptile volant et un reptile terrestre que les évolutionnistes prétendent être l'ancêtre le plus proche de ces créatures. Les différences entre les trois sont très grandes.
Lo scheletro del pipistrello
più antico che si conosca
(Icaronycteris) dell’Eocene.
Un moderno toporagno
che somiglia da vicino
agli antichi insettivori
che si afferma essere gli
antenati dei pipistrelli.
Il pipistrello più antico che si conosca e quello che gli evoluzionisti affermano essere l'antenato più prossimo. Notare la grandissima differenza tra il pipistrello e il suo cosiddetto antenato.
La ritenzione idrica si può vedere anche in piccoli dettagli. Per esempio, la balena nutre il cucciolo con una forma di latte concentrato simile al formaggio. Questo latte contiene dieci volte più grasso rispetto al latte umano. C'è una serie di ragioni chimiche per cui questo latte è così ricco di grasso. Quando la giovane balena digerisce il latte, si rilascia acqua. In questo modo la madre soddisfa i bisogni idrici della giovane balena con perdite minime di acqua.

Lo scheletro di
Plesiosaurus più
antico che si conosca
Scheletro di Araeoscelis, un rettile del Permiano inferiore.
l Pleisaurus, il rettile marino più antico che si conosca e il suo parente terrestre più prossimo, secondo gli evoluzionisti. Non c’è alcuna somiglianza tra i due.

Un tipico esempio delle balene più antiche che si conoscano, Zygorhiza kochi, dell’Eocene
Gli immaginari antenati della balena sonooggetto di dibattito tra le autorità evoluzioniste ma alcune di essehanno deciso per l’Ambulocetus. Alato c’è un‘Ambulocetus,un tipico tetrapodo
Un‘antica balena e quello che gli evoluzionisti affermano essere il suo antenato più prossimo. Notare che non c’è alcuna somiglianza tra di loro. Anche il miglior candidato che gli evoluzionisti hanno scelto come antenato delle balene non ha niente a che fare con loro.
2- Vista e comunicazione: gli occhi di delfini e balene consentono loro di avere una vista acuta in diversi ambienti. Hanno una vista perfetta anche in acqua. Tuttavia, la maggior parte degli esseri viventi, compreso l’uomo, vede poco al di fuori del suo ambiente naturale.
Gli occhi dei mammiferi terrestri e marini sono elaborati in modo stupefacente. Sulla terra, gli occhi affrontano una serie di potenziali pericoli. Ecco perché gli occhi dei mammiferi terrestri hanno palpebre che li proteggono. Nell’oceano, la più grande minaccia agli occhi viene dall’alto livello di sale e dalla pressione delle correnti. Per evitare il contatto diretto con le correnti, gli occhi sono situati ai lati della testa. Inoltre uno strato duro protegge gli occhi di creature che nuotano a grandi profondità. Gli occhi dei mammiferi marini sono dotati di elaborate funzioni che consentono di vedere in profondità dove c’è poca luce. Per esempio, il cristallino è di forma perfettamente circolare mentre nella retina i bastoncelli (le cellule sensibili alla luce) sono più numerosi dei coni (le cellule sensibili ai colori e ai dettagli). Inoltre gli occhi dei cetacei contengono anche uno strato di fosforo che li aiuta a vedere particolarmente bene al buio.

Un Halitherium, un‘antico lamantino dell’Oligocene.
L’irace, che è considerato il più prossimo antenato terrestre dei mammiferi acquatici sirenidi che comprendono anche i lamantini.
Un lamantino e quello che gli evoluzionisti chiamano il suo antenato terrestre più prossimo.

Scheletro di una foca moderna, virtualmente identica alle più antiche foche che si conoscano, del Miocene.
Cynodictis gregarius, il mammifero carnivoro abitante sulla terra che gli evoluzionisti credono sia stato il più prossimo antenato della foca.
Tipico scheletro di foca e quello che gli evoluzionisti credono sia il suo antenato terrestre più prossimo. Ancora una volta non c’è alcuna somiglianza tra i due.

Tutti i mammiferi marini confutano le immaginarie serie evolutive in cui si tenta di collocarli. La conclusione che emerge è che forme di vita diverse comparvero improvvisamente nel passato, indipendentemente l’una dall’altra. Questa è la prova scientifica del fatto che tutti gli esseri viventi furono creati.
La vista, comunque, non è la modalità sensoriale più importante per i mammiferi marini. Essi fanno affidamento sull’udito più di quanto di norma accadae nel caso dei mammiferi terrestri. La luce è essenziale per la vista, mentre l‘udito non ne ha bisogno. Molte balene e delfini cacciano in profondità dove è completamente buio, per mezzo di un meccanismo a sonar di cui dispongono. Le balene dentate, in particolare, “vedono” attraverso le onde sonore. Proprio come accade con le onde luminose nel sistema visivo, le onde sonore vengono messe a fuoco e analizzate e poi interpretate dal cervello. Questo dà al cetaceo informazioni accurate riguardo alla forma, alla dimensione, alla velocità e alla posizione dell’oggetto che ha di fronte. Questo sistema sonico è estremamente sensibile – per esempio un delfino può percepire una persona che si tuffa nell’acqua. Le onde sonore si usano anche per stabilire la direzione e per la comunicazione. Per esempio, due balene a centinaia di chilometri di distanza possono comunicare attraverso il suono.
Il problema del modo in cui questi animali producono suoni che consentono loro di stabilire la direzione o comunicare è ancora in gran parte irrisolto. Per quanto ne sappiamo, una particolare funzione del corpo dei delfini merita particolare attenzione: il cranio dell’animale, cioè, è isolato dal suono, una caratteristica che protegge il cervello da un continuo e intenso bombardamento sonoro.

L'existence des mammifères marins différents tels que les dauphins et les baleines ne prouve pas l'évolution mais la création.
Consideriamo il problema. È possibile che queste stupefacenti caratteristiche dei mammiferi marini siano giunte all’esistenza per mezzo della selezione naturale e delle mutazioni?. Quale mutazione avrebbe potuto avere come risultato il corpo dei delfini che possiede un sistema di sonar e un cervello isolato dal suono? Quale tipo di mutazione avrebbe potuto consentire ai suoi occhi di vedere nell’acqua buia? Quale mutazione avrebbe potuto portare al meccanismo che consente l’uso più economico dell’acqua?
Non c’è fine a tali domande e l’evoluzione non ha risposte da dare. Al contrario la teoria dell’evoluzione risponde con una storia incredibile. Consideriamo tutte le coincidenze che questa storia comporta nel caso dei mammiferi marini. Per prima cosa, i pesci erano appena venuti all’esistenza nell’acqua. Poi, per puro caso, fecero il passaggio verso la terra. Dopo di ciò si evolvettero sulla terra in rettili e mammiferi, ancora una volta solo per caso. Alla fine accadde semplicemente che alcune di queste creature tornarono in acqua dove, per caso, acquisirono tutte le caratteristiche di cui avevano bisogno per sopravvivere in tale ambiente.
La teoria dell’evoluzione può dimostrare anche una sola di queste fasi? Certamente no. Lungi dall’essere in grado di dimostrare l’affermazione nel suo insieme, la teoria dell’evoluzione è incapace di dimostrare come anche una sola di queste fase potrebbe essere avvenuta.
Conclusioni
Tutte le scoperte esaminate finora rivelano che le specie comparvero sulla terra all’improvviso e pienamente formate, senza alcun precedente processo evolutivo. Se è così, questa è la prova concreta che gli esseri viventi sono creati, come ha riconosciuto il biologo evoluzionista Douglas Futuyma. Ricordiamo che scrisse: “Se comparvero in uno stato di pieno sviluppo, devono in effetti essere stati creati da una qualche intelligenza onnipotente".144 Gli evoluzionisti, d’altro canto, cercano di interpretare la sequenza con cui gli esseri viventi comparvero sulla terra come prova dell’evoluzione. Siccome, però, tale processo evolutivo non ebbe mai luogo, questa sequenza può solo essere quella della creazione. I fossili rivelano che gli esseri viventi comparvero prima nel mare e poi sulla terra seguiti dalla comparsa dell’uomo.