Capitolo 12
Perchè Le Affermazioni Degli Evoluzionisti Sono Erronee
Nei capitoli precedenti, abbiamo constatato la mancanza di validità della teoria dell'evoluzione sulla base delle prove fornite dai fossili e dal punto di vista della biologia molecolare. In questo capitolo, analizzeremo una serie di fenomeni biologici e di concetti avanzati come prove teoretiche dagli evoluzionisti. Questi argomenti sono particolarmente importanti in quanto mostrano come non esista alcuna scoperta scientifica che avalli l'evoluzione, se non la distorsione e l'inganno.
Variazioni E Specie
Variazione, un termine usato in genetica, si riferisce a un evento genetico che provoca, in individui appartenenti a determinati tipi o specie, la presenza di caratteristiche diverse rispetto agli altri. Ad esempio, tutta la popolazione della Terra reca fondamentalmente le stesse informazioni genetiche, tuttavia alcuni presentano occhi allungati, altri i capelli rossi, alcuni hanno il naso lungo, altri sono di statura modesta, secondo il grado di variazione potenziale dell'informazione genetica.
Gli evoluzionisti affermano che le variazioni all'interno della specie rappresentano una prova a sostegno della loro teoria. Nondimeno, le variazioni non costituiscono una prova per l'evoluzione in quanto non sono altro che il risultato di differenti combinazioni di informazioni genetiche già esistenti a cui non aggiungono alcuna nuova caratteristica. La cosa importante per la teoria dell’evoluzione, però, è la questione di come informazioni del tutto nuove possano far esistere una specie del tutto nuova.
Le variazioni avvengono sempre entro i limiti dell'informazione genetica. Tale limite, in genetica, è detto "pool genetico", o "fondo comune di geni". Tutte le caratteristiche presenti nel pool genetico di una specie possono apparire in vari modi per variazione. Per esempio, a seguito della variazione, potrebbero comparire, in una specie rettile, varietà che presentano una coda più o meno lunga o zampe più corte, in quanto le informazioni per queste caratteristiche sono entrambe presenti nel pool genetico dei rettili. Nondimeno, le variazioni non trasformano questi ultimi in uccelli aggiungendovi le ali o le penne o mutando il loro metabolismo. Un tale cambiamento richiede un incremento delle informazioni genetiche degli esseri viventi, che non è assolutamente possibile nelle variazioni.
Darwin non era consapevole di questo fatto quando formulò la sua teoria. Egli credeva che non vi fossero limiti alle variazioni. In un articolo scritto nel 1844 afferma: "Molti autori sostengono che esista un limite alle variazioni in natura, benché io non sia in grado di scoprire un singolo fatto sul quale questa fede è fondata".143 Ne L'origine delle specie cita diversi esempi di variazioni come prove fondamentali a sostegno della sua teoria.
Ad esempio, secondo Darwin, gli allevatori di animali che accoppiavano diverse varietà di bovini per ottenere nuove varietà che producessero più latte, erano destinati infine a trasformarle in specie differenti. La nozione darwiniana di "variazione illimitata" è espressa con maggior chiarezza in un brano tratto da L'origine delle specie:
Non vedrei nessuna difficoltà che una razza di orsi per effetto della selezione naturale potesse diventare sempre più acquatica per struttura ed abitudini, con la bocca sempre più larga, fino a dar luogo ad un essere mostruoso come la balena.144
La ragione per cui Darwin ha citato un simile esempio deve essere ricondotta alla primitiva comprensione della scienza a lui contemporanea. Da allora, la scienza ha postulato il principio di "stabilità genetica" (omeostasi genetica) basata sui risultati degli esperimenti condotti su esseri viventi. Questo principio sostiene che, poiché tutti i tentativi di accoppiamento fatti per produrre nuove variazioni si sono rivelati inconcludenti, esistono rigide barriere tra le differenti specie di esseri viventi. Ciò significa che fu assolutamente impossibile per gli allevatori di animali trasformare il bestiame in specie differenti accoppiando diverse variazioni, come postulato da Darwin.
Le variazioni all’interno delle specie non implicano l’evoluzione
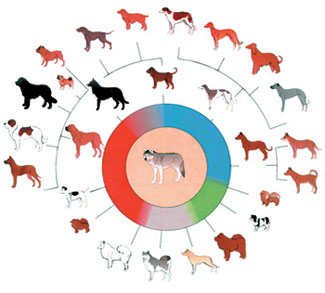
Nell’Origine delle specie, Darwin ha confuso due concetti distinti: le variazioni all'interno di una specie e l'apparizione di una nuova specie. Darwin osservò le varietà tra le varie razze di cani, per esempio, e immaginò che un giorno una di esse si sarebbe trasformata in una specie diversa. Ancora oggi gli evoluzionisti persistono nel tentare di presentare tali variazioni come “evoluzione”.
È una realtà scientifica, però, che le variazioni all’interno di una specie non sono evoluzione. Per esempio, indipendentemente dal numero razze di cani esistenti, questi rimarranno sempre una singola specie. Non ci sarà mai alcuna transizione da una specie distinta ad un’altra.
Norman Macbeth, che ha confutato il darwinismo nel suo libro Darwin Retried, scrive:
Il cuore del problema è se gli esseri viventi siano veramente variati illimitatamente... Le specie appaiono stabili. Tutti hanno sentito di allevatori delusi che hanno condotto il loro lavoro fino a un certo punto, per vedere poi gli animali o le piante tornare al punto da dove erano partiti. Nonostante gli strenui tentativi nel corso di due o tre secoli, non è mai stato possibile produrre una rosa blu o un tulipano nero.145
Luther Burbank, considerato l'allevatore più competente di tutti i tempi, esprime questo fatto quando afferma che "vi sono dei limiti allo sviluppo possibile, e questi obbediscono a una legge".146 Lo scienziato danese W. L. Johannsen riassume così la questione:
IL MITO DELLE BALENE CHE SI SONO EVOLUTE DAGLI ORSI

Nell'Origine della specie, Darwin affermò che le balene si sono evolute da orsi che tentavano di nuotare! Darwin supponeva erroneamente che le possibilità di variazione all'interno di una specie fossero illimitate. La scienza del ventesimo secolo ha dimostrato che questo scenario evoluzionistico è immaginario.
Le variazioni enfatizzate da Darwin e Wallace non possono essere selettivamente spinte oltre un certo limite, in quanto tale variabilità non contiene il segreto della "partenza indefinita".147
Allo stesso modo, i diversi fringuelli visti da Darwin sulle Isole Galápagos costituiscono un altro esempio della variazione, che non offre alcuna prova a sostegno dell’”evoluzione.” Osservazioni recenti hanno rivelato che i fringuelli non subiscono una variazione illimitata, come supponeva la teoria di Darwin. Inoltre, la maggior parte dei diversi tipi di fringuelli che Darwin pensava rappresentassero 14 specie in realtà si accoppiano tra di loro e questo vuol dire che sono variazioni all’interno della stessa specie. L’osservazione scientifica dimostra che i becchi dei fringuelli, che sono stati mitizzati in quasi tutte le fonti evoluzionistiche, sono in realtà un esempio di “variazione”; non costituiscono, quindi, una prova a favore della teoria dell’evoluzione. Ad esempio, Peter e Rosemary Grant, che hanno passato anni a osservare le varietà dei fringuelli nelle Isole Galápagos alla ricerca di prove per l’evoluzione darwinista, furono costretti a concludere che nessuna “evoluzione” che comportasse l’emergere di tratti nuovi sia mai avvenuto lì.148
La Resistenza Agli Antibiotici E L'immunità
Al DDT Non Sono Prove Dell'evoluzione
Uno dei concetti biologici che gli evoluzionisti si sforzano di presentare come prova per la loro teoria è la resistenza dei batteri agli antibiotici. Molte fonti evoluzioniste portano la resistenza agli antibiotici come "un esempio dello sviluppo degli esseri viventi mediante mutazioni vantaggiose". Un'affermazione simile viene fatta anche per gli insetti che hanno sviluppato immunità ad insetticidi come il DDT.
Tuttavia, gli evoluzionisti si sbagliano anche su questo argomento.
Gli antibiotici sono "molecole assassine" prodotte da microrganismi per combattere contro altri microrganismi. Il primo antibiotico fu la penicillina, scoperta da Alexander Fleming nel 1928. Fleming comprese che la muffa produceva una molecola che uccideva il batterio Staphilococcus, e questa scoperta segnò un punto di svolta nella medicina mondiale. Gli antibiotici derivati da microrganismi sono stati usati contro i batteri e i risultati sono stati positivi.
Presto venne fatta anche un'altra scoperta. Nel tempo, i batteri sviluppavano immunità agli antibiotici. Il meccanismo funziona così: una grossa parte dei batteri esposti agli antibiotici muore, ma altri, non influenzati dagli antibiotici, si replicano rapidamente e presto ricostituiscono l'intera popolazione. Così l'intera popolazione comincia a immunizzarsi agli antibiotici.
Gli evoluzionisti provano a presentare questo fatto come "l'evoluzione dei batteri mediante l'adattamento alle condizioni".
La verità, tuttavia, è molto diversa da questa interpretazione superficiale. Uno degli scienziati che ha fatto le ricerche più approfondite su questo argomento è il biofisico israeliano Lee Spetner, conosciuto anche per il suo libro Not by Chance (“Non per caso”) pubblicato nel 1997. Spetner sostiene che l'immunità dei batteri si produce mediante due diversi meccanismi, ma nessuno dei due costituisce una prova per la teoria dell'evoluzione. Questi due meccanismi sono:
1) il trasferimento di geni di resistenza già esistenti nei batteri.
2) la produzione di resistenza come risultato della perdita di dati genetici a causa della mutazione.
Il professor Spetner spiega il primo meccanismo in un articolo pubblicato nel 2001:
"Alcuni microrganismi sono dotati di geni che garantiscono resistenza a questi antibiotici. Questa resistenza può prendere la forma della degradazione della molecola antibiotica o della sua espulsione dalla cellula... Gli organismi che hanno questi geni possono trasferirli ad altri batteri rendendoli altrettanto resistenti. Sebbene il meccanismo di resistenza sia specifico a un antibiotico particolare, la maggior parte dei batteri patogeni sono... riusciti ad accumulare parecchi insiemi di geni che garantiscono loro resistenza ad una varietà di antibiotici".149
Spetner quindi continua dicendo che questa non è una "prova dell'evoluzione":
"L'acquisizione di resistenza agli antibiotici in questa maniera... non è del tipo che può servire come prototipo per le mutazioni necessarie a spiegare l'Evoluzione. I cambiamenti genetici che potrebbero illustrare quella teoria non soltanto devono aggiungere informazioni al genoma del batterio, ma devono aggiungere anche nuove informazioni al biocosmo. Il trasferimento orizzontale di geni non fa altro che diffondere dei geni che esistono già in alcune specie". 150
Quindi in questo caso non possiamo parlare affatto di evoluzione, perché non si produce alcuna nuova informazione genetica: le informazioni genetiche che già esistono sono semplicemente trasferite tra batteri.
Nemmeno il secondo tipo di immunità, che si produce come risultato della mutazione, è un esempio di evoluzione. Scrive Spetner:
"... Un microrganismo a volte può assumere una resistenza agli antibiotici mediante una sostituzione casuale di un singolo nucleotide... La streptomicina, che fu scoperta da Selman Waksman e Albert Schatz e studiata per la prima volta nel 1944, è uno degli antibiotici contro cui i batteri possono acquisire resistenza in questa maniera. Ma per quanto la mutazione che essi subiscono nel processo sia benefica per il microrganismo in presenza della streptomicina, non può servire come prototipo per il genere di mutazioni necessarie per la TND (Teoria Neodarwinista). Il tipo di mutazione che garantisce la resistenza alla streptomicina è evidente nel ribosoma e degrada il suo antagonista molecolare con la molecola antibiotica. Questo cambiamento nella superficie del ribosoma del microrganismo impedisce che la molecola di streptomicina si fissi e compia la sua funzione antibiotica. Ne risulta che questa degradazione è una perdita di specificità e quindi una perdita di informazioni. Il punto principale è che (l'Evoluzione) non si può ottenere mediante mutazioni di questo genere, non importa quante volte si riproducano. L'evoluzione non si può costruire accumulando mutazioni che degradano soltanto la specificità". 151
Gli evoluzionisti descrivono la resistenza dei batteri agli antibiotici come prova a favore dell’ evoluzione – ma in maniera ingannevole.
Per riassumere, una mutazione che si verifichi sul ribosoma di un batterio rende quel batterio resistente alla streptomicina. La ragione di ciò è la "decomposizione" del ribosoma mediante mutazione. Cioè, nessuna nuova informazione genetica viene aggiunta al batterio. Al contrario, la struttura del ribosoma è decomposta, in altre parole il batterio diventa "disabile" (inoltre, è stato scoperto che il ribosoma del batterio mutato è meno funzionale di quello di un batterio normale). Poiché questa "disabilità" evita che l'antibiotico si fissi sul ribosoma, si sviluppa una "resistenza antibiotica".
Infine, non c'è alcun esempio di mutazione che "sviluppi le informazioni genetiche".
La stessa situazione si rivela vera per l'immunità che gli insetti sviluppano nei confronti del DDT e di insetticidi simili. Nella maggior parte di questi casi, vengono utilizzati i geni immunitari che già esistono. Il biologo evoluzionista Francisco Ayala ammette questo fatto, e dice: "le varianti genetiche richieste per la resistenza ai diversi tipi di pesticidi apparentemente erano presenti in ciascuna delle popolazioni esposte a questi composti creati dall'uomo". 152
Altri esempi spiegati dalla mutazione, proprio come con la mutazione del ribosoma di cui sopra, sono i fenomeni che causano un "deficit di informazione genetica" negli insetti.
In questo caso non si può sostenere che il meccanismo dell'immunità nei batteri e negli insetti costituisca una prova per la teoria dell'evoluzione. Questo perché la teoria dell'evoluzione è basata sull'asserzione che gli esseri viventi si sviluppano attraverso mutazioni. Tuttavia, Spetner spiega che né l'immunità antibiotica né alcun altro fenomeno biologico indica un tale esempio di mutazione:
"Le mutazioni necessarie per la macro-evoluzione non sono mai state osservate. Nessuna mutazione casuale che possa rappresentare le mutazioni richieste dalla Teoria Neodarwinista e che sia stata esaminata al livello molecolare ha aggiunto alcuna informazione. La domanda che pongo è: le mutazioni osservate sono del genere che richiede la teoria come supporto? La risposta che ne risulta è: NO! 153
La Falsità Degli Organi Vestigiali
Per lungo tempo, il concetto di "organi vestigiali" appariva frequentemente nella letteratura evoluzionista come "prova" dell'evoluzione. Venne, infine, messo a riposo quando fu dimostrata la sua invalidità. Molti evoluzionisti, tuttavia, vi credono ancora e qualcuno tenta talvolta di proporre gli "organi rudimentali" come valida giustificazione della sua teoria.
Tale nozione fu avanzata per la prima volta un secolo fa. Come direbbero gli evoluzionisti, nei corpi di alcune creature, é esistito un certi numero di organi non funzionali. Questi organi erano stati ereditati dai progenitori ed erano gradualmente divenuti rudimentali a seguito del mancato uso.
L'intera ipotesi è alquanto anti-scientifica ed è interamente fondata su una conoscenza insufficiente. Questi "organi non funzionali" erano in realtà organi le cui "funzioni non erano ancora state scoperte". Ciò è confermato dal graduale, seppur sostanziale, decremento della lunga lista di organi rudimentali citati dagli evoluzionisti. S. R. Scadding, per quanto evoluzionista, confermò la validità di questo fatto in un articolo dal titolo "Possono gli organi rudimentali costituire una prova dell'evoluzione?" pubblicato sulla rivista Evolutionary Theory:
Dal momento che non è possibile identificare senza ambiguità strutture inutili e dal momento che la struttura dell'argomento utilizzata non è scientificamente valida, concludo che gli "organi vestigiali" non forniscono alcuna prova speciale per la teoria dell'evoluzione.154
La lista di organi rudimentali compilata dall'anatomista tedesco R. Wiedersheim nel 1895 comprese approssimativamente 100 organi, inclusi l'appendice e il coccige. Grazie ai progressi della scienza, venne scoperto che tutti gli organi inclusi nella lista di Wiedersheim avevano in realtà funzioni molto importanti. Ad esempio, si scoprì che l'appendice, che si supponeva fosse un organo rudimentale, era in realtà un organo linfatico che combatteva le infezioni del corpo. Questo fatto venne chiarito nel 1997: "Altri organi corporei e tessuti – il timo, il fegato, la milza, l'appendice, il midollo osseo e piccoli cumuli di tessuti linfatici come le tonsille nella gola e le placche di Peyer nell'intestino tenue– fanno parte del sistema linfatico. Anch'essi aiutano il corpo nella lotta contro le infezioni."155

Tutti gli esempi di organi vestigiali sono stati smentiti nel corso del tempo. Per esempio, ai giorni nostri, è stato dimostrato che la piega semicircolare dell'occhio, che fu citata nell‘Origine delle specie come struttura vestigiale, è pienamente funzionale, anche se la sua funzione era sconosciuta al tempo di Darwin. Questo organo lubrifica il globo oculare.
Venne, inoltre, scoperto che le tonsille, che erano state incluse nella lista, svolgevano un ruolo significativo nel proteggere la gola contro le infezioni, in particolare fino all'adolescenza. Si è anche compreso che il coccige, all'estremità della colonna vertebrale, sostiene le ossa attorno al bacino ed è il punto di convergenza di alcuni piccoli muscoli e perciò, non sarebbe stato possibile sedersi comodamente senza un coccige. Negli anni successivi, si scoprì che il timo stimolava il sistema immunitario nel corpo umano attivando le cellule T; che la ghiandola pineale era incaricata della secrezione di alcuni ormoni importanti; che la ghiandola tiroidea provvedeva alla solida crescita dei neonati e dei bambini; che la ghiandola pituitaria controllava il corretto funzionamento di molte ghiandole ormonali. Tutti questi erano un tempo considerati "organi rudimentali". Infine, è stato scoperto che la plica semilunare nell'occhio, che Darwin considerò un organo rudimentale, svolge in realtà il compito di pulire e lubrificare il globo oculare.
Quanto affermato dagli evoluzionisti riguardo agli organi rudimentali conteneva un grave errore logico. Come si è detto, essi affermavano che tali organi erano stati ereditati dai progenitori. Molti organi "rudimentali" non erano, tuttavia, presenti in quelle specie che si consideravano gli antenati degli esseri umani! Ad esempio, l'appendice non esiste in alcune specie di scimmie. Il noto biologo H. Enoch, che mise in dubbio la suddetta teoria, evidenziò l'errore logico in essa insito con le seguenti parole:
Le grandi scimmie possiedono un'appendice, mentre non è così per i loro parenti meno diretti, le scimmie inferiori; ma appare di nuovo tra i mammiferi ancora inferiori, quali l'opossum. Come possono spiegare tutto ciò gli evoluzionisti?156
Semplicemente, lo scenario degli organi rudimentali avanzato dagli evoluzionisti contiene una moltitudine di seri errori di logica ed è stato scientificamente confutato. Non esiste alcun organo rudimentale ereditato nel corpo umano, dal momento che gli esseri umani non si sono evoluti da altre creature per azione del caso ma furono creati nella loro attuale, completa e perfetta forma.
Il Mito Dell'omologia
Le somiglianze strutturali tra specie differenti sono dette in biologia "omologie". Gli evoluzionisti tentano di presentare tali somiglianze come prove della teoria in cui credono.
Darwin pensava che le creature con organi similari (omologhi) avessero una reciproca relazione evolutiva e che tali organi dovessero costituire l'eredità di un comune antenato. Secondo questo assunto, sia il piccione che l'aquila avevano ali; quindi, i piccioni, le aquile, e tutti gli altri uccelli provvisti di ali si supponeva che si fossero evoluti da un comune antenato.
L'omologia è una tesi ingannevole avanzata senza il supporto di alcuna prova, se non quella di un'apparente somiglianza fisica. Questo argomento non è mai stato verificato neppure da una singola scoperta concreta fin dai tempi di Darwin. In nessuna parte del mondo si è mai rinvenuto un fossile dell'immaginario antenato comune delle creature con strutture omologhe. Inoltre, ciò che segue appaleserà il fatto che l'omologia non fornisce alcuna prova all'evoluzione.
1. È possibile riscontrare organi omologhi in creature di phyla completamente differenti, tra cui gli evoluzionisti non sono in grado di stabilire alcuna sorta di relazioni evolutive.
2. I codici genetici di alcune creature che presentano organi omologhi sono completamente diversi tra loro.
3. Gli sviluppi embriologici di organi omologhi in creature diverse sono del tutto differenti.
Esaminiamo ora, singolarmente, ognuno di questi punti.
Aquile, pipistrelli e insetti hanno tutti le ali. Tuttavia, il solo fatto che posseggano organi simili non dimostra che si sono evoluti da un progenitore comune.
Organi Similari In Specie Viventi
Completamente Differenti
I gemelli mammiferi che sfidano l’omologia
DUE MAMMIFERI ESTINTI, SENZA RELAZIONE TRA DI LORO, CON DENTI GIGANTI
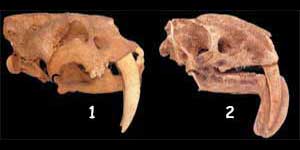
Un altro esempio di straordinaria somiglianza tra mammiferi “gemelli” placentati e marsupiali è quello tra i mammiferi estinti chiamati Smilodon (sotto) e Thylacosmilus (sopra), entrambi predatori con enormi denti anteriori. La grande somiglianza tra le strutture del cranio e dei denti di questi due mammiferi, tra cui non può essere stabilita alcuna relazione evolutiva, sovverte la visione omologica secondo cui strutture simili sono una prova a favore dell’evoluzione.
IL LUPO DI TASMANIA E IL SUO OMOLOGO NORDAMERICANO
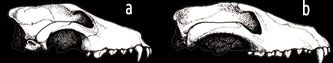

La presenza di specie “gemelle” tra mammiferi marsupiali e placentati dà un duro colpo alle affermazioni sull'omologia. Per esempio, il marsupiale lupo di Tasmania (sopra) e il lupo placentato trovato in Nord America, si somigliano in modo straordinario. Sopra si possono vedere i crani di questi due animali molto simili. Una così stretta somiglianza tra i due, che non si può immaginare abbiano una “relazione evolutiva”, invalida completamente le affermazioni dell’omologia.
a) Cranio di lupo nordamericano
b) Cranio di lupo della Tasmania
Esiste un certo numero di organi omologhi in comune tra gruppi differenti, tra i quali gli evoluzionisti non sono in grado di stabilire alcun tipo di relazione evolutiva. Le ali ne sono un esempio. Oltre agli uccelli, anche dei mammiferi come i pipistrelli ne sono provvisti, degli insetti e alcuni dinosauri, che sono dei rettili estinti. Neppure gli evoluzionisti stabiliscono una relazione o una parentela tra questi quattro differenti gruppi di animali.
Un altro esempio sorprendente è l'incredibile somiglianza e similarità strutturale osservata negli occhi di creature differenti. Ad esempio, il polpo e l'uomo sono due specie estremamente differenti, tra cui è impossibile qualsiasi tipo di relazione evolutiva, tuttavia gli occhi di entrambi sono molto simili in termini di struttura e funzione. Neppure gli evoluzionisti, per rendere conto della somiglianza degli occhi tra polpi ed esseri umani, pretendono che vi sia un antenato comune tra le due specie. Questo e numerosi altri esempi dimostrano che l'asserzione evoluzionista basata sulle somiglianze è completamente non scientifica.
In realtà, gli organi omologhi dovrebbero essere fonte di grave imbarazzo per gli evoluzionisti. Le confessioni del noto evoluzionista Frank Salisbury, che rivelano come creature estremamente differenti presentino occhi simili, sottolineano il vicolo cieco dell'omologia:
Anche qualcosa di talmente complesso come l'occhio è apparso parecchie volte; ad esempio nei calamari, nei vertebrati e negli artropodi. È già abbastanza difficile render conto dell'origine di ciò una sola volta, ma il pensiero di doverlo ripetere più volte, in accordo alla moderna teoria sintetica, mi provoca un giramento di testa.157
Ci sono molte creature che, pur avendo una costituzione fisica molto simile, non permettono alcuna pretesa di parentela evolutiva. Due grandi categorie di mammiferi, i placentati e i marsupiali, ne sono un esempio. Gli evoluzionisti ritengono che la distinzione tra di loro sia sorta quando i mammiferi sono apparsi per la prima volta e che ciascun gruppo abbia vissuto la propria storia evolutiva in maniera totalmente indipendente dall’altro. Ma è interessante notare che ci sono delle “paia” tra i placentati e i marsupiali che sono molto simili. I biologi statunitensi, Dean Kenyon e Percival Davis, commentano nella maniera seguente:
Secondo la teoria darwiniana, i modelli dei lupi, dei gatti, degli scoiattoli, dei ricci e dei topi si sono evoluti ciascuno due volte: una volta tra i mammiferi placentati e poi, in maniera del tutto indipendente, tra i marsupiali. Questo implica la straordinaria pretesa che un processo casuale e non diretto di mutazione e di selezione naturale sia, in qualche modo, capitato sulle stesse caratteristiche varie volte in organismi molto lontani tra di loro.158


In termini di struttura, gli occhi degli esseri umani e quelli dei polpi sono molto simili. Comunque, il fatto che le due specie abbiano organi simili non implica che si siano evoluti da un progenitore comune. Neppure gli evoluzionisti cercano di spiegare la somiglianza degli occhi di polpo e uomo ipotizzando un progenitore comune.
Le somiglianze straordinarie e organi simili come questi, che i biologi evoluzionisti non possono accettare come esempi di “omologia”, indicano che non ci sono prove per la tesi dell’evoluzione da un antenato comune. Quale potrebbe, in tal caso, essere la spiegazione scientifica di strutture simili negli esseri viventi? La risposta a questa domanda fu data prima che la teoria dell’evoluzione di Darwin dominasse il mondo della scienza. Gli scienziati come Carl Linnaeus, che per primo sistematizzò gli esseri viventi secondo le loro strutture simili, e Richard Owen, consideravano queste strutture come esempi di una creazione “comune”. In altre parole, gli organi simili (oppure, oggi, i geni simili) si ritiene che siano tali perché sono stati creati per servire a uno scopo particolare, non perché siano evoluti per caso da un antenato comune.
Le scoperte scientifiche moderne mostrano che la pretesa di un “comune antenato”, avanzata riguardo a simili organi, è incorretta, e che l’unica spiegazione possibile è una creazione comune, confermando così ancora una volta che gli esseri viventi sono stati creati da Dio.
L'impasse Genetica Ed Embriologica Dell'omologia
Affinché la teoria evoluzionista sull'omologia possa essere presa sul serio, gli organi simili (omologhi) nelle differenti creature dovrebbero presentare un simile (omologo) codice del DNA. Tuttavia, non è così. Gli organi simili sono governati, in genere, da codici genetici (DNA) molto diversi. Inoltre, codici genetici simili nel DNA di creature differenti sono spesso associati a organi completamente differenti.
Michael Denton, un professore australiano di biochimica, descrive nel suo libro Evolution: A Theory in Crisis il vicolo cieco che incontra l'interpretazione evoluzionista dell'omologia: "le strutture omologhe sono spesso contraddistinte da sistemi genetici non omologhi e il concetto di omologia può essere raramente esteso all'embriologia."159
Un famoso esempio di questo argomento è la “struttura scheletrica a cinque dita” dei quadrupedi, menzionata in quasi tutti i libri di testo evoluzionistici. I quadrupedi, cioè i vertebrati che abitano sulla terraferma, hanno cinque dita sugli arti anteriori e posteriori. Anche se queste non hanno sempre l’aspetto delle cinque dita come noi le conosciamo, sono tutti considerati pentadattili a causa della struttura ossea. Gli arti anteriori e posteriori di una rana, di una lucertola, di uno scoiattolo e di una scimmia hanno tutti la stessa struttura. Anche le strutture degli uccelli e dei pipistrelli sono conformi a questo disegno di base.
Gli evoluzionisti sostengono che tutti gli esseri viventi discendano da un antenato comune, e hanno a lungo citato l’arto pentadattilo come prova di ciò. Questa pretesa è stata citata in tutte le fonti fondamentali di biologia per tutto il Novecento come una prova forte dell’evoluzione. Le scoperte genetiche degli anni Ottanta hanno confutato questa affermazione evoluzionistica. Ci si rese conto che le strutture pentadattiliche di diverse creature erano controllate da geni completamente diversi. Il biologo evoluzionista William Fix descrive il crollo della tesi evoluzionista sul pentadattilismo come segue:
I libri di testo più vecchi sull’evoluzione danno molta importanza all’idea di omologia, indicando le evidenti somiglianze tra gli scheletri degli arti di animali diversi. Così il modello “pentadattilico” degli arti si trova nel braccio di un uomo, nell’ala di un uccello e nella pinna di una balena, e si ritiene che ciò indichi una loro comune origine. Purtroppo, non è così. Si sa oggi che gli organi omologhi sono prodotti da complessi di geni totalmente diversi nelle diverse specie. Il concetto di omologia in termini di geni simili, trasmessi da un antenato comune, è crollato.160
Inoltre, perché la tesi evoluzionista sull’omologia possa essere presa sul serio, dovrebbero essere paralleli i periodi di sviluppo embriologico di strutture simili, cioè gli stadi di sviluppo nell’uovo o nel ventre materno. In realtà, questi periodi embriologici per strutture simili sono assai diversi tra di loro in ogni creatura vivente.
Per concludere, si può affermare che la ricerca genetica e embriologica ha definitivamente confutato la definizione di Darwin, secondo cui il concetto di omologia sarebbe una "prova dell'evoluzione degli esseri viventi da un comune antenato." A questo riguardo, la scienza ha provato per l'ennesima volta la falsità della tesi darwinista.
Invalidità Dell'omologia Molecolare
La pretesa degli evoluzionisti che l'omologia costituisca una prova dell'evoluzione è invalidata non solo al livello morfologico, ma anche a quello molecolare. Essi affermano infatti che i codici del DNA o le strutture proteiche corrispondenti di differenti specie viventi siamo simili e che tale carattere sia una prova della loro evoluzione da un comune progenitore.

Professor Michael Denton: “L'evoluzione è una teoria in crisi”.
In realtà, però, i risultati delle comparazioni molecolari non operano a favore della teoria dell’evoluzione. Esistono enormi differenze molecolari tra creature che sembrano molto simili e imparentate. Ad esempio, la proteina citocroma-C, una delle proteine vitali per la respirazione, risulta incredibilmente diversa tra esseri viventi appartenenti alla stessa classe. Sulla base delle ricerche condotte in questo campo, la differenza tra due specie di rettili è maggiore di quella tra un uccello e un pesce o un pesce e un mammifero. Un altro studio ha mostrato che la differenza molecolare tra alcuni uccelli è superiore a quella tra alcuni uccelli e i mammiferi. È stato inoltre scoperto che la differenza molecolare tra batteri che appaiono molto simili è maggiore di quella tra mammiferi e anfibi o insetti.161 Simili confronti sono stati condotti nei casi di emoglobina, mioglobina, ormoni e geni con analoghi risultati.162
A proposito di queste e di altre scoperte nel campo della biologia molecolare, Michael Denton ha commentato:
A livello molecolare ogni classe è unica, isolata e disgiunta da passaggi intermedi. Così, le molecole, come i fossili, non hanno fornito gli elusivi passaggi intermedi tanto a lungo cercati dalla biologia evolutiva... A livello molecolare, nessun organismo è "ancestrale" o "primitivo" se comparato ai suoi parenti... Non c'è dubbio che se questa prova molecolare fosse stata disponibile un secolo fa... l'idea di evoluzione organica non sarebbe mai stata accettata.163
L'"Albero Della Vita" Sta Crollando
Negli anni Novanta, la ricerca sul codice genetico degli esseri viventi ha rafforzato il dilemma che la teoria dell’evoluzione già fronteggiava a questo proposito. In tali esperimenti, a differenza dei primi confronti che erano limitati alle sequenze di proteine, furono messe a confronto delle sequenze di "RNA ribosomico" (rRNA). Da queste scoperte, gli scienziati evoluzionisti hanno tentato di ricostruire un “albero evolutivo”. Tuttavia, furono delusi dai risultati. Secondo un articolo del 1999, dei biologi francesi Hervé Philippe e Patrick Forterre, "con sempre più sequenze disponibili, è emerso che la maggior parte delle filogenie proteiche si contraddicono tra di loro, come avviene con l’albero del rRNA”. 164
Oltre ai confronti di rRNA, anche le sequenze di DNA nei geni degli esseri viventi the DNA sono state comparate, ma i risultati sono stati opposti a quelli dell’ “albero della vita” presupposto dall’evoluzione. I biologi molecolari James A. Lake, Ravi Jain e Maria C. Rivera concludono in proposito in un articolo del 1999:
I ricercatori hanno cominciato con l’analizzare un gruppo di geni provenienti da organismi diversi e hanno osservato che le parentele tra di loro sono in contraddizione con l’albero evolutivo della vita ricavato dalla sola analisi del rRNA.165
Né i confronti che sono stati eseguiti per le proteine, né quelli del rRNA o dei geni, confermano le premesse della teoria dell’evoluzione. Carl Woese, un autorevole biologo dell’Università dell’Illinois, ammette così che il concetto di “filogenia” ha perso ogni significato di fronte alle scoperte molecolari:
Nessuna filogenia coerente di organismo è emersa dalle numerose singole filogenie proteiche prodotte finora. Si possono notare incongruenze filogenetiche ovunque nell’albero universale, dalle radici alle ramificazioni principali all’interno e tra i vari gruppi, fino alla formazione dei raggruppamenti principali stessi. 166
Il fatto che i risultati dei confronti molecolari non depongano a favore, ma semmai si oppongano, alla teoria dell’evoluzione, è ammesso anche nell’articolo intitolato "Is it Time to Uproot the Tree of Life?" pubblicato su Science nel 1999. Questo articolo di Elizabeth Pennisi afferma che le analisi genetiche e i confronti portati avanti dai biologi darwinisti allo scopo di far luce sull’“albero della vita” hanno portato di fatto a risultati diametralmente opposti, e prosegue dicendo che “i nuovi dati stanno intorbidendo il quadro evoluzionistico”:
Un anno fa, dei biologi che osservavano genomi recentemente sequenziali di più di una dozzina di microrganismi, pensavano che questi dati potessero confermare le linee essenziali della storia della vita primordiale finora accettate. Ma ciò che hanno visto li ha lasciati confusi. I confronti dei genomi disponibili non solo non hanno chiarito il quadro di come si siano evoluti i principali raggruppamenti della vita, ma l’hanno complicato. E oggi, che abbiamo in mano altre otto nuove sequenze microbiche, la situazione è diventata ancora più confusa.... Molti biologi evoluzionisti avevano pensato di poter approssimativamente vedere i primordi dei tre regni della natura… quando le sequenze complete del DNA hanno aperto la strada alla comparazione di altri tipi di geni, i ricercatori si aspettavano di dover semplicemente aggiungere dei dettagli a quell’albero. Ma “niente potrebbe essere più lontano dalla verità”, dice Claire Fraser, a capo del TIGR (The Institute for Genomic Research) a Rockville, Maryland. Invece, i confronti hanno portato molte varianti dell’albero della vita che differiscono da quelle dell’albero dei rRNA e che sono anche in conflitto tra di loro...167
In breve, man mano che la biologia molecolare progredisce, il concetto di omologia perde sempre più terreno. I confronti che sono stati eseguiti tra proteine, rRNA e geni rivelano che creature che erano ritenute molto prossime secondo la teoria dell’evoluzione, sono in realtà completamente differenti l’una dall’altra. Uno studio del 1996 che includeva 88 sequenze di proteine ha associato i conigli ai primati invece che ai roditori; un’analisi del 1998 di 13 geni in 19 specie animali ha piazzato i ricci di mare tra i cordati; e un altro studio del 1998 basato su 12 proteine ha posto le mucche più vicino alle balene che ai cavalli. Il biologo molecolare Jonathan Wells nel 2000 riassume la situazione in questo modo:
Le incongruenze tra alberi basati su molecole diverse, e i bizzarri alberi che risultano da alcune analisi molecolari, hanno spinto ora la filogenia molecolare in una crisi.168
“La filogenia molecolare” sta fronteggiando una crisi – il che vuol dire che anche la teoria dell’evoluzione sta fronteggiando una crisi (la filogenia si riferisce alle cosiddette “relazioni familiari” tra vari esseri viventi ed è la base ipotetica della teoria dell’evoluzione). Una volta di più, la scienza attacca alla radice la tesi secondo cui gli esseri viventi si sono evoluti gli uni dagli altri, dimostrando che tutti i gruppi viventi sono stati creati separatamente.
Il Mito Della Ricapitolazione Embriologica

Haeckel era un evoluzionista ancora più fervente di Darwin da molti punti di vista. Per questo motivo, non esitò a distorcere i dati scientifici e a inventare varie falsificazioni.
Ciò che una volta era chiamato "teoria della ricapitolazione" è stato da lungo tempo rimosso dalla letteratura scientifica, sebbene venga ancora presentato come una realtà da alcune pubblicazioni evoluzioniste. Il termine "ricapitolazione" è un condensato del detto "l'ontogenesi ricapitola la filogenesi" proposto dal biologo evoluzionista Ernst Haeckel alla fine del Ottocento.
La teoria avanzata da Haeckel postula che gli embrioni viventi ripercorrano il processo evolutivo dei loro progenitori. Egli teorizzò che nel corso del suo sviluppo nell'utero materno, l'embrione umano dapprima mostri le caratteristiche di un pesce, poi di un rettile ed infine quelle umane.
Da allora è stato provato che questa teoria non è altro che una fantasia. È ormai noto che le "branchie" che ipoteticamente appaiono nei primi stadi dell'embrione umano sono in realtà le fasi iniziali del condotto uditivo medio, della paratiroide e del timo. La parte dell’embrione che venne paragonata al "sacco vitellino" si è rivelata una sacca che produce sangue per l'infante. La parte che è stata identificata come una "coda" da Haeckel e dai suoi successori è in realtà la spina dorsale, la quale rassomiglia ad una coda solo perché prende forma prima delle gambe.
Questi fatti sono universalmente noti nel mondo scientifico e sono accettati anche dagli stessi evoluzionisti. George Gaylord Simpson, uno dei fondatori del neodarwinismo, scrive:
Haeckel travisò il principio evolutivo coinvolto. È ora fermamente stabilito che l'ontogenesi non ripete la filogenesi.169
In un articolo pubblicato su American Scientist si legge:
La legge biogenetica è sicuramente morta stecchita. È stata infine esorcizzata dai manuali di biologia nel corso degli anni Cinquanta. Come oggetto di serie indagini si estinse durante gli anni Venti...170
Un altro interessante aspetto della "ricapitolazione" fu Ernst Haeckel stesso, un ciarlatano che falsificò i suoi disegni al fine di promuovere la dottrina da lui avanzata. Le falsificazioni di Haeckel pretesero di mostrare che gli embrioni dei pesci e dell'uomo erano simili. Quando venne scoperto, si difese accusando anche altri evoluzionisti di aver commesso simili colpe:
Dopo questa compromettente confessione di "falsificazione" io dovrei considerarmi condannato e annichilito, se non avessi la consolazione di vedere accanto a me sul banco degli imputati centinaia di colleghi colpevoli, tra cui molti dei più fidati osservatori e dei più stimati biologi. La grande maggioranza di tutti i diagrammi presenti nei migliori testi di biologia, nei trattati e nei giornali presenta lo stesso grado di "falsificazione", in quanto sono tutti inesatti e più o meno manipolati, schematizzati e costruiti.171
I fraudolenti disegni di Haeckel
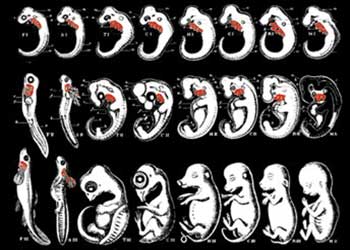
Questi disegni furono fabbricati da Haeckel per dimostrare le “similitudini” tra embrioni di esseri umani e di pesci. Confrontando il suo disegno con reali embrioni umani, si può vedere che egli ha deliberatamente omesso una larga parte dei veri organi. (Francis Hitching, The Neck of the Giraffe: Where Darwin Went Wrong, p. 205)

DISEGNO MANIPOLATO DISEGNO CORRETTO
Vi sono quindi "centinaia di colleghi colpevoli, tra cui molti dei più fidati osservatori e dei più stimati biologi" i cui testi sono pieni di pregiudizi, distorsioni e anche falsificazioni. Ciò è dovuto al fatto che tutti costoro si sono autocondizionati al fine di sostenere la causa della teoria evolutiva, sebbene non esista neppure un briciolo di prova scientifica in grado di avallarla.




