
Dans son livre The Blind Watchmaker (1986), l’athée Richard Dawkins se référa aux caractéristiques imparfaites dans la nature. Il apparut plus tard que son argument provenait de l’ignorance.
Autrefois Régnait Le Mythe Des Caractéristiques Imparfaites
Le professeur de zoologie de l’Université d’Oxford Richard Dawkins est l’un des plus célèbres évolutionnistes au monde aujourd’hui. Il est connu non pas pour ses travaux en zoologie mais pour sa défense ardente du darwinisme et de l’athéisme.

En 1986, il publia le livre The Blind Watchmaker (L’horloger aveugle) qui essaie de convaincre les lecteurs que les caractéristiques complexes des créatures vivantes étaient le résultat de la sélection naturelle. Ses tentatives reposaient pour la plupart sur des spéculations, des comparaisons défectueuses et des calculs faux que divers scientifiques et écrivains ont exposés en détail depuis.66
Les caractéristiques “imparfaites” ou “mauvaises” faisaient partie des arguments de Dawkins. Il affirmait que certaines structures chez les créatures vivantes étaient inutiles et qu’elles étaient donc imparfaites pour éliminer le principe d’une création parfaite. L’exemple principal qu’il fournit est celui de la rétine inversée de l’œil chez les vertébrés, y compris chez l’homme.
Une rétine inversée signifie que les photorécepteurs sont localisés à l’arrière de l’œil et non à l’avant, là où la lumière pénètre. Les terminaisons sensibles de ces cellules percevant la lumière font face à l’arrière et les nerfs rétiniens qui en ressortent forment une couche entre la lumière et les cellules. Ces nerfs convergent vers un point sur la rétine par lequel ils quittent l’œil. Parce qu’il n’y a pas de photorécepteurs à ce stade, c’est le “point aveugle” de l’œil, où il n’y a pas de vision.
Les darwinistes ont fait de cette inversion et de ce point aveugle des défauts, car selon eux, l’œil est issu de la sélection naturelle et il faut s’attendre par conséquent à de telles bizarreries. Richard Dawkins est encore une fois un fidèle partisan de cet argument. Dans The Blind Watchmaker (L’horloger aveugle), il écrivit:
N’importe quel ingénieur supposerait naturellement que les photocellules pointent vers la lumière, avec leurs fils partant en arrière vers le cerveau. Il rirait si on suggérait que les photocellules puissent pointer ailleurs que vers la lumière, avec leurs fils partant du côté le plus proche de la lumière. Pourtant, c’est précisément ce qu’il se passe dans l’œil de tous les vertébrés.67
Pourtant, Dawkins et ceux qui partagent son avis ont tort parce qu’ils ignorent l’anatomie et la physiologie de l’œil.

Michael Benton, professeur de biologie
Michael Denton de l’Université d’Otago, biologiste moléculaire et critique acerbe du darwinisme aujourd’hui, donne un compte-rendu détaillé de la question. Dans “The Inverted Retina: Maladaptation or Preadaptation?” (La rétine inversée: mal-adaptation ou pré-adaptation?), paru dans le magazine Origins and Design, il expliqua comme la rétine inversée passée pour imparfaite par Dawkins est en réalité créée de la manière la plus efficace pour l’œil du vertébré:
La prise en compte des grandes exigences en énergie des cellules des photorécepteurs dans la rétine du vertébré suggère qu’au lieu d’être un défi à la téléologie, la curieuse conception inversée de la rétine est peut-être l’unique solution au problème consistant à fournir aux cellules photoréceptrices hautement actives chez les vertébrés supérieurs de copieuses quantités d’oxygène et de nutriments.68

Pour comprendre ce point souligné par le professeur Denton mais passé inaperçu chez Dawkins, nous devons d’abord reconnaître que les cellules des photorécepteurs de la rétine ont besoin d’une quantité importante d’énergie et d’oxygène. Pendant que nos yeux ouverts perçoivent la lumière, il s’y produit des réactions chimiques très complexes toutes les secondes. Les photons, les plus petites particules de lumière, sont perçus par les cellules. Suite aux réactions chimiques entamées par les protons, la perception a lieu et est répétée à chaque instant. Cette réaction est si complexe et rapide que selon les termes de Denton, “la couche de photorécepteur a l’un des taux métaboliques les plus hauts des tissus connus”.69
Pour maintenir ce taux de métabolisme élevé, les cellules de la rétine ont besoin de beaucoup d’énergie. Les cellules de la rétine chez un être humain consomment 150% d’énergie de plus que les cellules rénales, trois fois plus que les cellules du cortex cérébral et six fois plus que les cellules composant le muscle cardiaque. Cette comparaison est établie sur la base de la couche de rétine entière; les photorécepteurs, soit moins de la moitié de cette couche, ont, quant à eux, besoin de plus d’énergie que l’ensemble de la couche. Dans son ouvrage encyclopédique, The Vertebrate Eye (L’œil chez le vertébré), G. L. Walls qualifie les photorécepteurs d’“avides” à la fois de nutriments et d’oxygène.70
Comment ces cellules qui nous permettent de voir parviennent-elles à pourvoir à leurs besoins nutritionnels et en oxygène?
A travers le sang bien-sûr, comme pour le reste du corps.
Mais alors d’où vient le sang?
A ce stade, nous pouvons comprendre pourquoi la rétine inversée est un signe parfait de la création. Juste à l’extérieur de la couche de rétine se trouve un important tissu de veines similaire à un filet. Denton écrit:
L’oxygène et les nutriments pour satisfaire l’appétit métabolique vorace des photorécepteurs sont fournis par un lit unique de capillaires, appelé choriocapillaires, correspondant à un réseau de grands capillaires aplatis qui forment une couche vasculaire riche située à la sortie immédiate des photorécepteurs, séparés d’eux uniquement par la couche de cellules épithéliales de la rétine (RPE) et une membrane spéciale – la membrane de Bruch – qui forment ensemble une barrière hautement sélective ne laissant passer dans la rétine que les métabolites et les nutriments nécessaires au fonctionnement de RPE et des photorécepteurs. Ces capillaires sont beaucoup plus grands que les capillaires standards, mesurant 18 à 50 microns de diamètre. Ce réseau unique de vaisseaux sanguins donne l’impression d’être spécialement adaptés pour fournir à la couche photoréceptrice de copieuses quantités de sang.71
Dans son livre, An Introduction to the Biology of Vision (Introduction à la biologie de la vision), le professeur James T. McIlwain écrit: “A cause des grands besoins métaboliques des photorécepteurs, l’œil semble avoir adopté une stratégie d’inonder la choroïde de sang afin d’assurer que l’approvisionnement ne soit jamais un problème.”72
C’est pour cette raison que les photorécepteurs sont inversés. Il y a clairement une explication derrière cet état. L’arrangement inversé de la rétine n’a rien de défectueux comme le proclama Dawkins, mais constitue une preuve de la création dans un but spécifique.
Dans un article pertinent, Denton examine si la rétine pouvait être constituée autrement. Sa conclusion est que cela n’était pas possible. Si l’on envisageait, comme le suggéra Dawkins, une rétine plate avec des cellules réceptrices face à la lumière, ces dernières s’éloigneraient des capillaires les nourrissant et se trouveraient privées d’une partie de l’oxygène et des nutriments dont elles ont besoin. Etendre les capillaires dans la rétine ne résoudrait pas le problème parce que cela produirait de nombreux points aveugles et réduirait par conséquent la capacité de l’œil à voir.
Denton commente:
Plus on s’intéresse de près à la rétine des vertébrés et plus il apparaît que chaque caractéristique y est nécessaire. Si nous cherchions à reconcevoir depuis les principes de base un œil capable de la plus haute résolution et de la plus haute sensibilité (capable de détecter un photon individuel de lumière) nous finirions par recréer l’œil du vertébré – complet avec une rétine inversée… 73
En résumé, les arguments de Dawkins et des autres évolutionnistes cherchant à faire croire que “la rétine chez les vertébrés est défectueuse” découlent de l’ignorance. Leurs conclusions ont été annulées par les investigations plus informées sur la minutie des créatures vivantes. Mais il ne s’agit pas là du seul argument issu de l’ignorance au cours de l’histoire du darwinisme. Voyons ce qu’il en est du mythe des vestiges d’organes.
Le Mythe Des Vestiges D’organes
Vous avez certainement dû lire que chez l’homme l’appendice et le coccyx, ou os de la queue, sont des vestiges d’organes qui autrefois avaient d’importantes fonctions chez nos prétendus ancêtres de l’évolution, mais qui au fil du temps perdirent leurs fonctions.
Nombreux sont ceux qui en entendirent parler parce que depuis Darwin, le mythe des vestiges d’organes est la matière à propagande préférée des évolutionnistes.
Le mythe fut lancé avec la mention dans L’origine des espèces d’organes dont les fonctions se perdirent ou se réduisirent. Darwin qualifiait ces organes de “rudimentaire” et les comparait “aux lettres qui, conservées dans l’orthographe d’un mot, bien qu’inutiles pour sa prononciation, servent à en retracer l’origine et la filiation”.74 En 1895, l’anatomiste allemand R. Wiedersheim proposa une liste d’une centaine de vestiges d’organes humains, comprenant notamment l’appendice et le coccyx.
Etant donné le niveau primitif de la science du 19ème siècle, l’appendice était considéré comme un organe inutile et donc comme un vestige.
Mais comme le reste des idées darwinistes, ce mythe put se développer en raison du niveau peu sophistiqué de la science à l’époque. Grâce aux progrès effectués en science, on découvrit lentement que les organes perçus par Darwin et ses disciples comme des vestiges avaient de réelles fonctions. On découvrit ainsi que les organes de la liste dressée par Wiedersheim étaient très utiles au corps. Au fur et à mesure des découvertes, cette liste se raccourcit sensiblement. Il fut démontré par exemple que l’appendice avait un rôle très important dans le système lymphatique luttant contre les germes pénétrant dans le corps. Un article intitulé “Examples of Bad Design Gone Bad” (Les exemples de conception défectueuse tournant mal), se référant à plusieurs ouvrages de base en anatomie, explique:
L’examen au microscope de l’appendice montre qu’il contient une quantité considérable de tissus lymphoïdes. Des agrégats similaires de tissu lymphoïde (tissus lymphoïdes associés à l’intestin–GALT) se produisent dans d’autres régions du système gastro-intestinal. Les GALT sont impliqués dans la capacité du corps à reconnaître les antigènes étrangers dans les matières ingérées. Ma propre recherche, en particulier, se concentre sur l’examen des fonctions immunologiques de l’intestin.
Des expériences menées sur des lapins montrent que l’appendicectomie néonatale affecte le développement de l’immunité muqueuse. Des études morphologiques et fonctionnelles de l’appendice du lapin indiquent qu’il représente probablement l’équivalent de la bourse de Fabricius chez les mammifères. La bourse joue un rôle essentiel dans le développement de l’immunité humorale chez les oiseaux. La similarité histologique et immuno-histochimique de l’appendice du lapin et de l’homme suggère que l’appendice humain a une fonction similaire à celle du lapin. L’appendice humain peut être spécialement important dans les premières années parce qu’il achève son développement peu après la naissance et ensuite régresse avec l’âge, ressemblant finalement à d’autres régions du GALT tels que les plaques de Peyer dans l’intestin grêle. Ces études récentes démontrent que l’appendice humain n’est pas un vestige d’organes, comme cela fut avancé à l’origine.75
En bref, la raison pour laquelle l’appendice fut considéré comme un vestige vient du dogmatisme de Darwin et de ses partisans, et avec la contribution du niveau peu sophistiqué de la science à l’époque. Avec les microscopes rudimentaires dont ils disposaient, ils ne pouvaient pas observer les tissus lymphatiques de l’appendice. Puisqu’ils ne pouvaient pas en comprendre la structure, ils la jugèrent inutile et l’ajoutèrent à leur liste des vestiges d’organes inutiles. Une fois de plus, le darwinisme fut encouragé par l’absence de précision de la science au 19ème siècle.
Il en fut de même pour tous les autres organes contenus dans la liste de Wiedersheim. Les amygdales étaient assimilées à des vestiges d’organes alors qu’on découvrit leur contribution dans la protection de la gorge contre les infections, particulièrement avant l’âge adulte. L’os de la queue, situé à la base de la colonne vertébrale s’avéra, quant à lui, soutenir les os autour du pelvis et par conséquent, en son absence il serait difficile de s’asseoir confortablement. On comprit alors également que cet os constitue le point à partir duquel tous les organes et les muscles de la région pelvienne sont maintenus ensemble.
Il fut, par ailleurs, démontré que le thymus active les cellules T et met en route le système immunitaire du corps. La glande pinéale est responsable de la sécrétion des hormones essentielles telles que la mélatonine contrôlant la production de l’hormone lutéinisante. La glande de la thyroïde assure le développement équilibré de l’enfant et la mise en place du taux métabolique dans le corps. La glande pituitaire contribue au bon fonctionnement de glandes hormonales telles que la thyroïde, les glandes surrénales et les glandes reproductives. Elle contrôle également le développement du squelette.
Le pli semi-lunaire au coin de l’œil dont Darwin avait fait un vestige aide en réalité à garder l’œil propre et à le lubrifier.
Aujourd’hui, il ne fait plus de doute que les organes qualifiés autrefois de vestiges ont tous des fonctions bien définies. Dans leur livre Vestigial Organs are Fully Fonctionnal (Les vestiges d’organes sont pleinement fonctionnels), les docteurs Jerry Bergman et George Howe reprirent ce point en détail.
Il est désormais reconnu que le mythe des vestiges d’organes souscrits par tant d’évolutionnistes fait partie des arguments basés sur l’ignorance. Dans “Do Vestigial Organs Provide Evidence for Evolution?” (Les vestiges d’organes sont-ils une preuve de l’évolution), un article paru dans le magazine Evolutionary Theory, le biologiste évolutionniste S.R. Scadding écrit:
A mesure que nos connaissances ont augmenté, la liste des vestiges de structures s’est réduite… Puisqu’il n’est pas possible d’identifier sans ambiguïté des structures inutiles, et puisque la structure de l’argument utilisé n’est scientifiquement pas valable, je conclus que les organes vestigiaux n’apportent aucune preuve particulière à la théorie de l’évolution.76
Même s’il fallut plus d’un siècle et demi pour le faire admettre aux évolutionnistes, un autre mythe du darwinisme vient de s’évaporer.
Le Pouce Du Panda

Stephen Jay Gould
Au début de ce chapitre, il a été question de la rétine des vertébrés censément imparfaite selon Richard Dawkins. Stephen J. Gould, paléontologue évolutionniste à l’Université d’Harvard, défendait des idées similaires. Avant son décès en 2002, il faisait partie des principaux évolutionnistes aux Etats-Unis.
Comme Dawkins, Gould s’intéressa aux caractéristiques “défectueuses”, en l’occurrence le pouce du panda.
Contrairement à l’homme, le panda n’a pas de pouce opposable distinct de ses quatre autres doigts lui facilitant la préhension des objets. Ses cinq doigts s’étendent côte à côte. Mais en plus de ces cinq doigts parallèles, le panda présente une projection sur le poignet, appelée “os sésamoïde radial”. Le panda l’utilise parfois comme un doigt, c’est pourquoi les biologistes lui ont donné le nom de pouce du panda.
Gould prétendit que cet os dans la main du panda n’était pas fonctionnel. Il était si convaincu de l’importance de sa thèse qu’en 1980, il publia un livre sur le sujet.
Or, comme pour Dawkins, la théorie de Gould sur les caractéristiques défectueuses était fausse. L’erreur de Gould provenait du fait qu’il comparait la main du panda à celle de l’homme, supposant ainsi que le pouce du panda avait la même fonction. Paul Nelson fit le commentaire suivant:
Bien que le pouce du panda soit peut-être sous-optimal pour de nombreuses tâches (comme la frappe), il semble adapté pour ce qui serait sa fonction usuelle, le décorticage des bambous.77

Dans son livre paru en 1980, The Panda’s Thumb, Gould suggéra que la main de l’animal présentait un défaut. Or les découvertes scientifiques prouvèrent qu’au contraire le pouce du panda était hautement fonctionnel.
Les auteurs de The Giant Pandas of Wolong (Les pandas géants de Wolong) avancent:
Le panda peut manipuler les pousses de bambous avec une grande précision, en les saisissant comme avec un forceps dans les fentes imberbes reliant le coussinet du premier doigt au pseudo-pouce… Lorsqu’on observe un panda manger des feuilles, on est toujours impressionné par sa dextérité. Les pattes avant et la bouche travaillent ensemble avec une grande précision et une grande économie de mouvements… 78
Une étude publiée en 1999 par le magazine Nature montrait que dans son environnement naturel, le pouce du panda était extrêmement utile. Ce projet commun mené par quatre chercheurs japonais recourait à la tomographie et aux techniques d’imagerie à résonance magnétique. Il fut conclu que le pouce du panda est “l’un des systèmes de manipulation les plus extraordinaires”79 qui soient chez les mammifères. Le commentaire suivant est tiré du même article intitulé “Role of the Giant Panda’s Pseudo-thumb” (Le rôle du pseudo-pouce du panda géant):
Nous avons montré que la main du panda géant présente un mécanisme de préhension bien plus précis que les modèles morphologiques précédents le suggèrent.80
En résumé, les théories avancées par les évolutionnistes pendant les 150 dernières années concernant les organes vestigiaux et les caractéristiques biologiques imparfaites se sont avérées fausses grâce à des recherches plus avancées sur les structures en question.
Les évolutionnistes ne peuvent guère expliquer les origines d’une quelconque structure biologique dans la nature. Tous leurs arguments s’opposant au concept de création ont été infirmés.
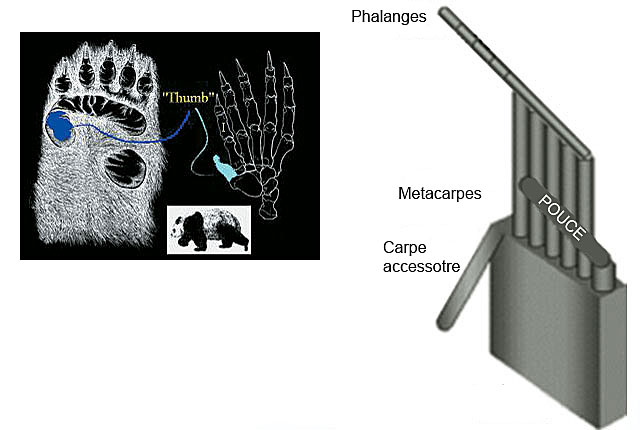
Le pouce du panda est un excellent exemple de conception.
Afin de nier la création, les évolutionnistes cherchent des défauts et des incohérences dans la nature. La thèse de Gould à propos du pouce du panda en est un exemple. Gould commit une erreur, puisque cet os de pouce n’est pas un défaut étant donné qu’il facilite le mouvement et prévient les déchirures des tendons.
Une étude publiée dans le magazine Nature le 28 janvier 1999 montra que le pouce du panda était très utile dans l’habitat naturel de l’animal. Cette étude conjointe menée par quatre chercheurs japonais à l’aide de la tomographie informatique et l’imagerie à résonnance magnétique conclut que le pouce du panda était “l’un des systèmes de manipulation les plus extraordinaires” chez les mammifères. (Endo H., Yamagiwa D., Hayashi Y.H., Koie H., Yamaya Y. et Kimura J. 1999 Nature 397 : 209-310). Ci-dessus, un modèle schématique de la structure de la main du panda préparé par les experts ayant mené cette étude.
C’est pourquoi nous pouvons affirmer qu’autrefois existait le darwinisme, dont les défenseurs prétendaient que les êtres vivants possédaient de nombreux organes vestigiaux défectueux.
Cette théorie est désormais discréditée par les découvertes scientifiques.

NOTES
66- Dawkins'in "kör saatçi" tezinin çürülmesi için bkz. Lee Spetner, Not By Chance: Shattering the Modern Theory of Evolution, Judaica Press, 1997; Michael J. Behe, Darwin's Black Box: The Biochemical Challange to Evolution, The Free Press, 1996; Phillip E. Johson, Darwin on Trial, 199, 2nd.ed., InterVarsity Press, 1993 ![]()
67- Richard Dawkins, The Blind Watchmaker, London: Penguin Books,1986; s.93-94 ![]()
68- Michael Denton, "The Inverted Retina: Maladaptation or Pre-adaptation?", Origins & Design, 19:2, Issue 37, 1999 ![]()
69- Michael Denton, "The Inverted Retina: Maladaptation or Pre-adaptation?", Origins & Design, 19:2, Issue 37, 1999 ![]()
70- Walls, G.L. (1963). The Vertebrate Eye. New York: Hafner Publishing Company; s.652 ![]()
71- Michael Denton, "The Inverted Retina: Maladaptation or Pre-adaptation?", Origins & Design, 19:2, Issue 37, 1999 ![]()
72- McIlwain, T.J. (1996). An Introduction to the Biology of Vision. Cambridge: Cambridge University Press; s. 14 ![]()
73- Michael Denton, “The Inverted Retina: Maladaptation or Pre-adaptation?”, Origins & Design, 19:2, Issue 37, 1999 ![]()
74- Charles Darwin, The Origin of Species, III. ed. Chapter 13: Mutual Affinities of Organic Beings: Morphology: Embryology: Rudimentary Organs ![]()
75- http://www.cerrah.net/apandist.htm ![]()
76- www.geocities.com/CapeCanaveral/Lab/6562/evolution/designgonebad.html ![]()
77- S. R. Scadding, "Do 'Vestigial Organs' Provide Evidence for Evolution?", Evolutionary Theory, Cilt 5, Mayıs 1981, s. 173 ![]()
78- Paul A. Nelson, "Jettison the Arguments, or the Rule? The Place of Darwinian Theological Themata in Evolutionary Reasoning", Access Research Network, 1988, http://www.arn.org/docs/nelson/pn_jettison.htm ![]()
79- George Schaller, H. Jinchu, P. Wenshi, and Z. Jing, The Giant Pandas of Wolong, Chicago: University of Chicago Press, 1986, s.4, 58 ![]()
80- Endo, H., Yamagiwa, D., Hayashi, Y. H., Koie, H., Yamaya, Y, and Kimura, J., Nature, vol. 397, 1999, ss. 309-310 ![]()
PARTAGER
Téléchargements
- Introduction
- L’écroulement des mythes du Darwinisme et la définition correcte de la science
- Autrefois la vie était supposée être simple
- Autrefois les archives fossiles étaient supposées prouver l’évolution
- Autrefois fut lancée la quête du chaînon manquant
- Autrefois on ne disposait pas de l’information biologique
- Autrefois les preuves embryologiques étaient supposées prouver l’évolution
- Autrefois régnait le mythe des caractéristiques imparfaites
- Autrefois existait le mythe de ı’adn poubelle
- Autrefois, l’origine des espèces était supposée se trouver dans la spéciation
- Autrefois existait le scénario de “la série de chevaux”
- Autrefois il y avait l’histoire des mites poivrées
- Récemment encore, il était question d’histoires sur le dino-oiseau
- Conclusion
LES SUJETS
CavourLe VaticanLe socialismeL'illuminationLa Révolution FrançaiseConvertiSabbatéenLe jacobinismeLe Média MaçonniqueLe Sionisme PolitiqueLes Turcs JeunesLa Commitée de l'Union et du ProgrèsAbdulhamidAnti-NaziL'Organisation Sioniste MondialeLes Lois de NurembergMussoliniLa Guerre Mondiale IAdolf EichmannGoyimLa dynastie de RothschildThink-ThankCFRRockefellerLa Guerre FroideStalinLa Révolution d'OctobreL'union SoviétiqueBilderbergViêt-NamAIPACLobbyfoireSud-estgrècenouvel ordre mondialla mer rougegéopolitiquevétéranimpôtdouane2023antilopetaureauConseil islamique de l'eurasieprix de nobelhôpitalinstitution de sécurité nationalAli BabacanTurgut OzalassasinatGaffar OkkanMuhsin YazıcıoğluRosetta Nebulaastronomierose fleursermon de damasarme nucléaireIyad AllawiNautilussous-marincellule de plongéeAli BardakoglutélévisiontelevoleCanakkale