The Sudden Appearance of Major Animal Groups
Evolutionists claim that fish evolved from such invertebrate sea creatures as pikaia; amphibians and modern-day fish from some ancestral fish; reptiles from amphibians, birds and mammals from separate groups of reptiles—and finally that human beings and present-day apes evolved from a common ancestor.
In order to demonstrate the scientific veracity of these claims, they need to be able to show fossils of transitional creatures that represent a clear turning point in the development of these species. As already made clear, however, there is not the slightest trace of these imaginary creatures. For that reason, evolutionists persist in their biased interpretations of some fossils, suggesting that these represent transitional forms. Yet these obligatory transitional links are the subject of much controversy, even among evolutionists themselves. Not one so-called transitional link has ever been unconditionally accepted, because these are not actually transitional links at all. However, since evolutionists are obliged to come up with some such progression, they interpret some of the fossils they have found as intermediate forms.
Gareth Nelson of the American Museum of Natural History has this to say about evolutionists’ arbitrary selection of evolutionary ancestors:
We’ve got to have some ancestors. We’ll pick those. Why? “Because we know they have to be there, and these are the best candidates.” That’s by and large the way it has worked. I am not exaggerating.25
This chapter will be examining the scientific evidence for how living things emerged on Earth independently and did not, as evolutionists claim, evolve from one another.
The True Origin of Fish
According to evolutionist claims, invertebrates were the ancestors of the first vertebrate fish. Yet how did these creatures—with only a hard shell in some cases, and no bones or spinal column—turn into vertebrates? This is a question evolutionists are unable to answer, and for which they can find no evidence. That is because the living things in question would have to have undergone such enormous changes that while their shells were becoming vestigial on the outside, skeletons were forming on the inside. In order for such a huge transformation to take place, there would have to be an enormous number of intermediate forms between the two groups. The fact is, however, that there exists not a single fossil that evolutionists can present as a transitional form between invertebrates and vertebrates.
The theory of evolution assumes that first chordates such as pikaia gradually turned into fish. This claim was frequently repeated by evolutionists in the 1990s. Stephen Jay Gould, one of the most prominent contemporary exponents of Darwinism, declared pikaia to be “the ancestor of us all.” His claim rested on the assumption that vertebrates did not exist during the Cambrian Period. The portrayal of pikaia, the oldest known chordate—in other words, an animal with a central nervous system, which emerged during the Cambrian Period—as the ancestor of the fishes identified during later periods, appeared entirely compatible with the fossil record.
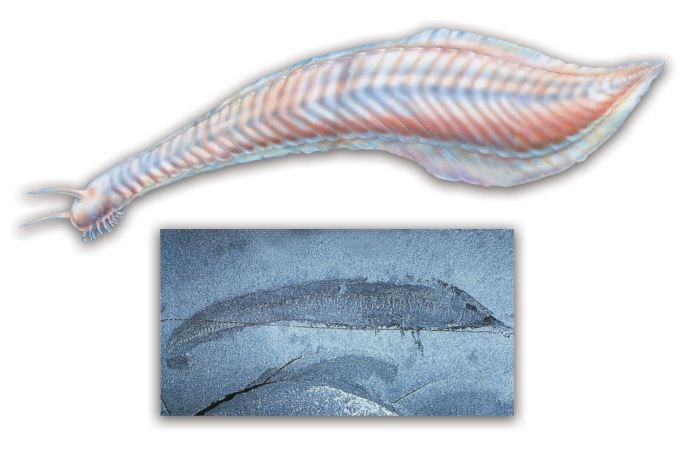 |
Evolutionists maintain that the creature known as the pikaia is the ancestor of fish. The fact is, however, that fish supposedly descended from the pikaia have been shown to have lived at the same time as it, during the Cambrian Period.
A pikaia fossil (bottom) |
A discovery made in China in 1999, however, undermined this evolutionist thesis concerning the Cambrian Period, proving that there were indeed fish living at the same time as pikaia, the supposed ancestor of all fish.
Paleontologists excavating in the Yunnan region found 530-million-year-old fossil fish. In his report, "Waking up to the Dawn of Vertebrates", the well-known paleontologist Richard Monastersky made the following statement regarding these two separate fish species, Haikouichthys ercaicunensis and Myllokunmingia fengjiaoa:
Paleontologists have long regarded vertebrates as latecomers who straggled into evolutionary history after much of the initial sound and fury had fizzled. Chinese paleontologists, however, have discovered fossils of two fish that push the origin of vertebrates back to the riotous biological bash when almost all other animal groups emerged in the geologic record. Preserved in 530-million-year-old rocks from Yunnan province, the paper clip-size impressions record the earliest known fish, which predate the next-oldest vertebrates by at least 30 million years.26
With this realization that vertebrates already existed in the Cambrian Period, the theory of evolution’s “tree of life” metaphor lost all credibility. All the basic categories of life, vertebrates included, emerged during the same geological period, so evidently that there can be no question of “evolution” from some common ancestor.
 |
A new discovery in 1999 revealed the existence of two fish species that lived in the Cambrian Period.
Top: Haikouichthys ercaicunensis
Bottom: Myllokunmingia fengjiaoa |
The fact that fish appeared at exactly the same time as all other complex groups shows that they did not evolve from any other species, but were created. Indeed, after the Cambrian Period, all the different categories of fish appear suddenly in the fossil record, with no ancestors behind them.
From Fish to Amphibians
According to evolutionists, the ancestor of land-dwelling life forms was some species of fish. They suggest that this imaginary creature, whose remains they have so far been unable to unearth, was forced to live in shallow, muddy water because of drought. For this reason, its descendants’ fins evolved into feet and their gills into lungs. They developed kidneys to remove their bodily wastes, and their skin acquired features preventing it from losing moisture—and that the first amphibian was the result.
Unless a fish underwent all these changes and more, it would be unable to live on land, and would die within a few minutes at most.
Evolutionists nominate three different fish species as the ancestors of amphibians. One of these is the famous “living fossil,” the coelacanth. Due to the thickness of its fins and certain of its bony structures, this species was for years portrayed as the ancestor of amphibians. In 1938, however, when a living specimen was caught in the Indian Ocean, it was realized that all the speculations that evolutionists had been engaging in was incorrect. Some 200 other living coelacanths were caught in the years that followed. When these were studied, it was clear that this species’ soft tissues has no resemblance to amphibians’, that they were not about to emerge onto land and that they swam in very deep waters, not shallow ones. (For further details, see the chapter on "False Transitional Forms.")
 |
A modern-day living coelacanth.
When the first live coelacanth, which evolutionists had for years suggested was a transitional form, was caught in the Indian Ocean in 1938, it was realized that this fish was not a transitional form at all. Contrary to what evolutionists had claimed, it was not a creature preparing for the transition from sea to land. Indeed, it actually lived in rather deep waters. Neither was any structure found in its fins to resemble feet, again contrary to what evolutionists had maintained. |
Since the coelacanth is now “extinct” as an the ancestor of amphibians, the great majority of evolutionists today propose a replacement: a group of fish from the Rhipidistia family. The fins of these fish contain bones and tissue as thick as those in the Coelacanth. Due to these different structures, evolutionists claim that legs first began to appear in this species. In fact, however, these structures bear no similarity to the front or hind legs of land-dwelling creatures. Moreover, just as with the coelacanth, these creatures’ fins are loosely bound to their skeletal muscles, but not connected to the backbone in such a way as to support their bodies’ weight. In other words, these fishes’ fins display no features resembling the legs of land-dwelling animals. Furthermore, the oldest known fossil amphibians have a pelvis and shoulders that are broad and powerful—features entirely absent in fish. No vestigial traces of a transition to such dynamic structures have been found in the so-called ancestors proposed by evolutionists.
 |
| The Australian lung fish, which evolutionists maintain is the ancestor of amphibians. Yet there is no resemblance between these creatures’ lungs and those of landdwelling animals. |
For the role of amphibian ancestor, evolutionists’ third candidates are lungfish (from the Dipnoi family). These species can breathe air on the surface as well as through their gills. However, the lung structure they possess has no similarities at all to the lungs of land-dwelling creatures.
These fishes’ skeletal structure is also very different to that of amphibians. For instance, there is no trace of legs in the fishes’ fin structure. Not only the backbone is very different, but so is the structure of the internal organs. These animals would need to undergo enormous changes in order to evolve into amphibians. As the pelvic bone formed, for instance, the gills would have to be replaced with lungs, and the ears and eyes would have to become capable of functioning in dry air.
 |
| According to the theory of evolution, land-dwelling animals evolved from fish. If this were true, then half-fish half-reptile creatures of the kind shown should be found in the fossil record. Yet there is no sign in the record that such creatures ever existed. |
No matter what species of fish evolutionists choose to regard as the so-called ancestor of amphibians, the number of changes necessary for a fish to develop into an amphibian is enormous. For that reason, there would have to be an outlandish number of transitional links between the two: strange half-finned/half-footed, half-gilled/half-lunged, half-kidneyed creatures must have existed, and would have had to number in the millions. However, not one of them has ever been encountered in the 100 million or so fossils from all over the world. There are complete fish, and there are complete amphibians in the fossil record, but no such transitional forms. This is a fact that refutes the theory of evolution, and is accepted even by evolutionists.
For example, Professor Robert Wesson of the Massachusetts Institute of Technology, describes how amphibians appear suddenly in the fossil record and how there is no evidence of any transition to them from fish:
The stages by which a fish gave rise to an amphibian are unknown. There are resemblances between the first amphibians and certain (rhipidistian) fish with bony fins, but the earliest land animals appear with four good limbs, shoulder and pelvic girdles, ribs, and distinct heads. . . . In a few million years, 320 million years ago, a dozen orders of amphibians suddenly appear in the record, none apparently ancestral to any other.27
 |
| Acanthostega: A creature that evolutionists suggest as an example of the transition from fish to amphibians. Yet it is not actually a transitional form at all. |
As Wesson makes clear, land-dwellers appear suddenly in the fossil record, with four healthy feet, shoulders, a ribcage and all the other features peculiar to them. No fossil has been found which can be claimed as these animals’ evolutionary ancestor, as is also stated by professor of Biology Keith Stewart Thomson emeritus Professor of Natural History at the University of Oxford:
While we still do not have any really intermediate fossil forms between fishes and tetrapods, we are free to argue vociferously about the identity of the group of fishes that must be the tetrapod ancestor. 28
From Amphibians to Reptiles
According to the Darwinist claim, reptiles such as crocodiles, lizards and snakes evolved from amphibians. But amphibians and reptiles possess very different properties in a wide range of respects.
One of the most distinctive differences between the two is their egg structures. Since amphibians lay their eggs in water, their eggs possess a structure necessary for development there. The eggs have a permeable, transparent membrane and a jelly-like structure. However, the structure of reptile eggs has been created to be suited to a dry terrestrial environment. The leathery shell of a reptile’s egg, also known as the amniotic egg, allows the passage of air, but not of water. In this way. the liquid needed by the developing embryo is preserved until the hatchling emerges from its egg.
 |  |
| One of the main differences between amphibians and reptiles is the structure of their eggs. Amphibians’ eggs are transparent and permeable, in a manner suited to an aquatic environment, whereas reptiles have shelled eggs suited to the land. | A Complete amphibtan (picture: 1)
Imaginary transitional forms (picture: 1-4)
A Complete reptile (picture: 5) No transitional form exists showing that reptiles evolved from amphibians. |
If amphibians were to lay eggs on land they would shortly desiccate, and the embryos inside would die. This is a difficulty that cannot be explained by the theory of evolution, which maintains that reptiles evolved gradually from amphibians. If life on land is to continue, then the amphibian’s egg must turn into an amniotic one within a single generation. No one can explain how this can be accomplished by natural selection and mutation, the two proposed mechanisms of evolution.
On the other hand, the fossil record strips the origin of reptiles of any evolutionary explanations. In an article titled “Problems of the Origins of Reptiles,” the well-known evolutionist paleontologist Robert L. Carroll accepts the truth of this:
Unfortunately, not a single specimen of an appropriate reptilian ancestor is known prior to the appearance of true reptiles. The absence of such ancestral forms leaves many problems of the amphibian-reptilian transition unanswered.29
Carroll, regarded as an authority on vertebrate paleontology, is forced to accept that “The early amniotes are sufficiently distinct from all Paleozoic amphibians that their specific ancestry has not been established.”30 The same thing is admitted by Stephen Jay Gould, who wrote, “No fossil amphibian seems clearly ancestral to the lineage of fully terrestrial vertebrates (reptiles, birds, and mammals).”31
 |
| When it was realized that Seymouria, which evolutionists maintained was the ancestor of reptiles, actually lived at the same time as them, that evolutionary claim had to be abandoned. |
The most important animal to be suggested so far as a reptilian ancestor is the amphibian species Seymouria. However, fossil reptiles have been discovered that were alive 30 million years before Seymouria’s first appearance on Earth, showing that this could not have been a transitional form. The oldest Seymouria fossils date back to the Sub-Permian Period, 280 million years ago. However, the two oldest known reptile species, Hylonomus and Paleothrys, were found in Sub-Pennsylvanian strata, which dates back to between 330 and 315 million years ago.32 It is therefore of course impossible for the ancestor of reptiles to have appear many millions of years after reptiles themselves.
In short, scientific findings show that reptiles did not appear on Earth as the result of a gradual development, as the theory of evolution maintains, but that they emerged suddenly—with no ancestors behind them.
The True Origin of Marine Reptiles
Marine reptiles are yet another group for whose origins evolutionists cannot account. In our time, sea turtles and sea snakes are members of this group. The most important extinct marine reptile is the Ichthyosaurus, which evolutionists suggest evolved from land-dwelling reptiles. But they are unable to explain how this might have taken place, however, and cannot provide any corroborating evidence from the fossil record.
Ichthyosaurs possessed the complex and unique features of species that live in oceans and deep water. Yet evolutionists maintain that some land-dwelling reptile adapted to life in deep, open waters as the result of chance.
 Ichthyosaur, an extinct sea reptile |
This is an impossible scenario. A. S. Romer, an expert on the natural history of vertebrates, states that a very long period of time would be necessary for the features peculiar to the Ichthyosaur to have emerged—for which reason the origin of these creatures must go back a very long way. He then accepts that no known Permian Period reptile could possibly be regarded as these creatures’ ancestor.33 This observation, made by Romer in the 1960s, is still valid today.
An article called “Rulers of the Jurassic Seas,” published in a special supplement to Scientific American magazine in April 2003, stated that Ichthyosaurs were suited not only to life on the coasts, but also to the ocean depths—for which reason they would have to undergo extreme adaptations in order to cross from the land to the sea, losing a great many terrestrial features and acquiring new ones for life in water.34 This, however, would require a very long time before the final animal actually emerged, via a tremendous number of transitional forms. Yet in the fossil record, there is no trace of such transitional forms that might be regarded as the ancestors of the Ichthyosaurs. Those fossils to date discovered are either of land-dwelling reptiles or marine ones.
It will be useful to compare certain features of Ichthyosaurs and land-dwelling reptiles to demonstrate how impossible it is for the latter to have evolved into the former:
• One of the main features which distinguish Ichthyosaurs from land-dwellers is the formers’ wide, flat flipper -like feet they used for swimming. Such flat feet do not appear in land-dwellers. Unlike the thin bones in many reptiles’ front feet, the bones in Ichthyosaurs’ front feet are short and wide.
• Moreover, the bones in their feet are all similar. In most four-footed animals, it is easy to distinguish the wrist bones from the palm bones. Of even greater importance, there is no flesh between the Ichthyosaurbones. They are squeezed very closely together, providing a hard, resistant surface. The fact that all its bones are contained within a single structure of tissue increases the firmness of the reptile’s feet. Interestingly, the same structure exists in present-day whales, dolphins, seals and turtles. Palette feet also increase their hydrodynamic efficiency, since their very shape reduces water resistance. If the toes were separated, this could not happen. The question of how Ichthyosaurs’ feet—or for that matter, the feet of sea turtles and marine mammals—came into being by evolution is an unanswered one. There is no evidence whatsoever of a transition to such a structure, from either fish fins or from the feet of land-dwelling reptiles.
• Even Scientific American accepts that there was no gradual, progressive transition to a flipper of this kind, and goes on to say:
Indeed, analyses of ichthyosaur limbs reveal a complex evolutionary process in which digits were lost, added and divided.35
As we have seen, the alleged evolution of Ichthyosaurs’ palette feet shows no constant development of the type that evolutionists expect. Like all evolutionist publications, however, Scientific American ignores this fact and with classic demagoguery, makes the following statement so that its readers too should ignore the truth:
Needless to say, evolution does not always follow a continuous, directional path from one trait to another.36
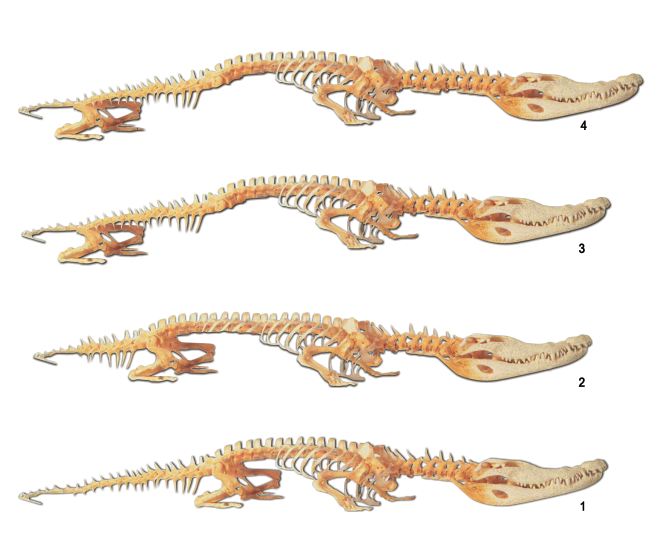 |
| A fossil crocodile with a reptile form (picture 1)
There is no trace of these imaginary transitional forms (pictures 2-4) in the fossil record. Another difference between reptiles and Ichthyosaurs is the number of vertebrae toward the front of their bodies. Reptiles have 20 vertebrae, and Ichthyosaurs up to 40. That being so, there should be fossils of transitional forms in the evolutionary process with, say, 25, 35 or 38 vertebrae. Yet there is in fact no sign of such fossils. |
When evolutionists fail to find their expectations confirmed, they attempt to save the theory by making statements of that sort. The fact is, though, that findings from the fossil record clearly show that no evolution ever occured.
• Another difference between reptiles and Ichthyosaurs is the number of vertebrae in the front part of their spines. There are only up to 20 vertebrae in the front of reptile spines, but up to 40 in the Ichthyosaur. In other words, during the so-called evolutionary process, the mutations that supposedly affected these creatures must have given them an extra 20 vertebrae, in addition to all other changes. Again, as you might expect, there are no fossil traces of reptiles that demonstrate a transitional number of vertebrae (creatures with 25, 30 or 35 vertebrae, for instance).
• Animals that hunt in the open sea can only find very limited amounts of prey. They need to swim in a very productive, energy-efficient manner. A tail-like fin is ideal for such swimming. The animal waves its fin like a stern oar to increase its mobility. Ichthyosaurs possessed just such a fin. Yet there is no fossil trace of any biological structure that could represent a forerunner of such a fin.

Ichthyosaurs were marine reptiles specially designed to live in deep waters. There is absolutely no evidence of these creatures’ imaginary evolutionary ancestors in the fossil record. Left: a 200-million- year-old Ichthyosaur fossil.
As we have seen, Ichthyosaurs possess exceedingly complex features specially created to let them live in deep ocean waters. To enjoy those specifications, a land-dweller must undergo many beneficial mutations. Yet random chance cannot change every feature of a living creature in a planned manner and compatible with its specific environment. Random coincidences cannot change a land-dwelling creature’s toes, its vertebrae, the structure of its eyes and the way it swims, nor the kind of designs necessary to enable it to live in deep water.
Coincidence lacks the intelligence and intention to do this. Indeed, the fossil record demonstrates that these reptiles came into being not through phased coincidences, but in a single moment, with all their complex and particular structures.
In their book Evolution of the Vertebrates, Colbert and Morales say this about these creatures’ origin:
The Ichthyosaurs, in many respects the most highly specialized of the marine reptiles, appeared in early Triassic times. Their advent into the geologic history of the reptiles was sudden and dramatic; there are no clues in pre-Triassic sediments as to the possible ancestors of the Ichthyosaurs . . . The basic problem of Ichthyosaur relationships is that no conclusive evidence can be found for linking these reptiles with any other reptilian order.37
The vertebrate paleontologist Chris McGowan describes how Ichthyosaurs appear suddenly in the fossil record, with no evolutionary ancestors preceding them:
I suggested that ichthyosaurs had just dropped out of the sky. The embarrassing fact is that we have not yet found the ancestor of the Ichthyosaurs. This has not prevented paleontologist from speculating, though, and most reptilian groups, at one time or another have been proposed as possible ichthyosaur ancestors.38
As McGowan—an evolutionist—courageously admits, the lack of evidence represents no obstacle to evolutionists producing fictitious ancestors for marine reptiles. Even so, evolutionist speculation is insufficient to conceal the manifest truth that, like all other creatures, marine reptiles were created. For that reason, no fossils belonging to their ancestors are to be found anywhere in the fossil record.
The True Origin of Mammals
According to the theory of evolution, some reptiles evolved into birds and others into mammals. But there are distinct and considerable differences between mammals and reptiles. Reptiles are cold-blooded, reproduce by laying leathery-shelled eggs, and their bodies are covered in scales. All reptiles have seven bones in their lower jaws, but only one bone in their ears.
 |
| Pictures 1-2: The crocodile a complete reptile
Imaginary transitional forms (pictures 3-5)
Picture 6:A Souirrel a complete mammal Yet there is not a single example of this and other imaginary transitional forms ! Picture 7: A complete tortoise, of which many fossils exist
Picture 8: There is no trace of this or similar imaginary transitional forms
Picture 9: A complete rabbit,of which fossils do exist There are fossils of rabbits, tortoises, lizards and squirrels in the fossil record, but not a single fossil of the half-mammal, half-reptile creatures of which evolutionists dream. |
Mammals, on the other hand, are warm-blooded, give birth to their young, and have milk glands and fur. They have a single lower jawbone and three bones in their ears, known to anatomists as the hammer, anvil and stirrup. If mammals, with their exceedingly complex and interconnected systems and structures, really did evolve from reptiles as the result of mutations, then in the fossil record there should be a large number of species reflecting that transition. We would expect to find remains of creatures possessing semi-developed milk glands, with scales mutating into fur, some legs further extended and others still short like a reptile’s, and similar half-formed, incomplete features. Yet not a single fossil of that type has ever been found. If creatures meeting that description had ever existed, their fossil remains would have survived.

A fossil of a creature belonging to the class Therapsida. Evolutionists depict these as the ancestors of mammals. Yet that claim is not scientific.
In addition, evolutionists suggest that the numerous kinds of mammals, from horses to human beings and from squirrels to elephants, all evolved from reptiles. They also claim that the emergence of mammals lasted some 100 million years. In order for such a large number of species to have made their appearance over in such a long period of time, then they should have left millions of transitional- form fossils. Yet not one of these expected transitional forms has ever been encountered in the fossil record. Evolutionists merely point to a group of fossils belonging to the group Therapsida, which are known as “mammal-like reptiles” as examples of transitional forms. But as you’ll see in the chapter on “False Transitional Forms,” their claims are invalid.
These mammal-like reptiles indicated as having been the ancestors of mammals are all extinct. And these creatures too emerge suddenly in the fossil record, and disappear just as suddenly.
Their being extinct gives evolutionists the opportunity to speculate about them however they wish, as they used to do with the coelacanth. But, looking at constructing resemblances between species on the basis of only a few bones is not a reliable method. Some evolutionists err in thinking that animals with similar skeletons also possess similar soft tissues. Michael Denton makes the following comment on this error by evolutionists:
Further, there is always the possibility that groups, such as the mammal-like reptiles which have left no living representative, might have possessed features in their soft biology completely different from any known reptile or mammal which would eliminate them completely as potential mammalian ancestors, just as the discovery of the living coelacanth revealed features in its soft anatomy which were unexpected and cast doubt on the ancestral status of its rhipidistian relatives.39
After studying the skulls brains of so-called mammal-like reptiles, scientists concluded that these creatures did not possess mammalian features, but entirely resembled reptiles. Mammals are distinguished from all reptiles (including mammal-like reptiles) by the size of their brains:
Similar considerations cloud the status of other classic intermediate groups such as the mammal-like reptiles, a group of extinct reptiles in which the morphology of the skull and jaw was very close to the mammalian condition. The possibility that the mammal-like reptiles were completely reptilian in terms of their anatomy and physiology cannot be excluded. The only evidence we have regarding their soft biology is their cranial endocasts, and these suggest that, as far as their central nervous systems were concerned, they were entirely reptilian. Jerison, who has probably had more experience studying the cranial endocasts of fossil species than any other authority in the field, comments on the mammal-like reptile brains: “. . . these animals had brains of typical lower vertebrate size. . .” Since their endocasts were all very near the volume of the expected brain sizes and since the endocasts present maximum limits on their brain sizes, the mammal-like reptiles could not have had brains that approached a mammalian size. . . . The mammal-like reptiles, in short, were reptilian and not mammalian with respect to the evolution of their brains. . . . The earliest mammal for which there is reasonable evidence, Triconodon of the upper Jurassic period, was apparently already at or near the level of living “primitive” mammals such as the insectivores or the Virginia opossum.40
Actually, mammal-like reptiles were described as such merely on the basis of similarities in the jaw joints. The fact is, however, that one single feature is not sufficient to allow such a definition.41
Studies on these creatures have concluded that they have nothing in common with mammals. In 1973, for example, Morganucodon was described by Dr. K. A. Kermack and other researchers from the University of London as Cynodont, a so-called transitional form with advance reptile features. A number of Morganucodon fragments were found in both Wales and China, showing that the same “evolutionary” stages had been undergone at more or less the same time, in two completely different parts of the world, divided from one another by thousands of miles—which is impossible. The researchers stated that from the point of view of their jawbones, Morganucodon and the earlier discovered Kuehneotherium, were both fully reptilian.42
 |
| Roger Lewin |
Another problem regarding these creatures, claimed to constitute transitional forms between reptiles and mammals, has to do with timing. These mammal-like reptiles emerge not at the end of the great Age of Reptiles, but at its beginning. This, according to the imaginary evolutionary tree, means that they appeared 100 million years too early.
In his evolutionist article “The Reptiles that Became Mammals,” published in New Scientist magazine, Tom Kemp admits that mammal-like reptiles appear suddenly in the fossil record:
As is now well known, most fossil species appear instantaneously in the fossil record persist for some millions of years virtually unchanged, only to disappear abruptly.43
All of this shows that the assumption that reptiles evolved into mammals has no scientific basis. The dilemma which forced the evolutionist paleontologist Roger Lewin to admit, “The transition to the first mammal . . . is still an enigma” still applies.44
On the other hand, the origin of the classes of mammals is also shrouded in darkness as regards the theory of evolution. As the evolutionist zoologist Eric Lombard writes in Evolution magazine:
Those searching for specific information useful in constructing phylogenies of mammalian taxa will be disappointed.45
As with all other groups, the origin of mammals is in not the least compatible with the theory of evolution.
The True Origin of Marine Mammals
Just like land-dwelling mammals, whales and dolphins give birth, suckle their young, breathe through their lungs and are warm-blooded. The origin of this group known as sea mammals is one of the most difficult for evolutionists to explain. In most of their sources, they suggest that sea mammals’ ancestors lived on land and over a long period of time, evolved, in such a way as to adapt to life in a marine environment. According to this view, marine mammals—whose alleged ancestors, fish, are assumed to have undergone a transition from sea to the land—returned to the water as the result of further so-called evolutionary pressures. The fact is however, no paleontological evidence supports this theory, which also flies in the face of logic.
 |
| Evolutionists’ Claims of a “walking whale” Are Unscientific 1: Pakicetus(50 million years old)
2: Ambulocetus (49 million years old)
3:Kutchicetus (43-46 million years old)
4: Rodhocetus (46.5 million years old)
5: Dorudon(37 million years old)
6: Basilosaurus (37 million years old)
7: Pakicetus |
The theory of evolution’s claim regarding the origin of whales rests on a fossil sequence, in which a series of species are arranged in an imaginary sequence and then proposed as transitional forms of whales’ evolution.
According to evolutionists, the geological sequence followed by these creatures is as follows: Pakicetus (50 million years ago) > Ambulocetus (49 million years ago) > Rodhocetus (46.5 million years ago) > Procetus (45 million years ago) > Kutchicetus (43 to 46 million years ago) > Dorudon (37 million years ago) > Basilosaurus (37 million years ago) > and finally, Aetiocetus (24 to 26 million years ago).
But there are a number of deceptive aspects to this scheme. The most fundamental of these—the first two creatures in the plan, Pakicetus and Ambulocetus—were both, according to evolutionists, “walking whales.” Yet to describe these two land-dwelling mammals as whales is an illusory, even comic claim to make.
First, consider Pakicetus inachus.
 |
| 1: A drawing of ambulocetus
2: The Basilosaurus fossil shown in the imaginary drawing to the side is one of the largest known whales.
3: An Archaeoceti (an archaicor early whale) skullı |
Fossils of this extinct mammal first entered the equation in 1983. The discoverers of the fossil, Philip D. Gingerich and his colleagues, had no hesitation in declaring it to represent a “primitive whale,” though they had found only its skull.
But in fact, this fossil has absolutely nothing to connect it to whales. Its four-legged skeleton resembles that of present-day wolves. The fossil was discovered in a stratum containing iron oxide ore as well as fossils of such land-dwellers as snails, tortoises and crocodiles. In other words, it was part of the land, not of a one-time sea bed.
This four-legged land-dweller was declared to be a “primitive whale” merely for certain details in its teeth and ear bones! The fact is, though, these features are no evidence on which to build a relationship between Pakicetus and today’s whales. Even evolutionists admit that positing such theoretical relationships among living things, by taking anatomical similarities as a starting point, are usually exceedingly inaccurate. If the platypus—a billed, egg-laying mammal living in Australia—and ducks were both extinct, then evolutionists, employing the same logic and taking these species’ similar bills and eggs as their starting point, would declare the two to be related. Yet the platypus is a mammal, and ducks are birds, and the theory of evolution can establish no relationship between them. Similarly, Pakicetus inachus, which evolutionists declare to be a primitive whale, is a unique species with its own particular anatomical features. Even Carroll, a foremost authority on vertebrate paleontology, states that the Mesonychid family, in which Pakicetus should be included, “was the combination of these changes.” 46 Even prominent evolutionists such as Gould accept that such “mosaic creatures” cannot be regarded as transitional forms.
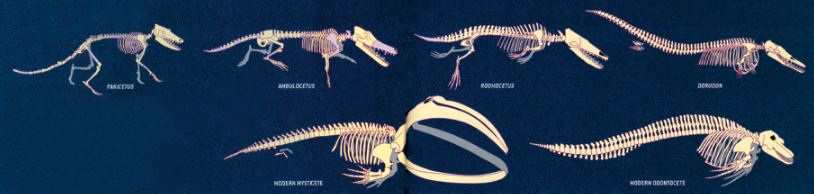 |
| Evolutionists’ Imaginary Whale Diagram When fossils of the creatures which evolutionists place here are examined, it is clear that there are enormous anatomical differences between them, but no transitional forms link them together. |
In an article titled “The Overselling of Whale Evolution,” the creationist writer Ashby L. Camp explains the invalidity of the claim that the Archaeocetae (whose Latin name means “archaic whales”)— part of the class Mesonychid, of which land mammals such as Pakicetus are members—are in fact whales:
The reason [why] evolutionists are confident that mesonychids gave rise to archaeocetes, despite the inability to identify any species in the actual lineage, is that known mesonychids and archaeocetes have some similarities. These similarities, however, are not sufficient to make the case for ancestry, especially in light of the vast differences. The subjective nature of such comparisons is evident from the fact so many groups of mammals and even reptiles have been suggested as ancestral to whales.47
In the whales’ imaginary evolutionary tree, the second extinct creature after Pakicetus is Ambulocetus [Latin for “walking whale’] natans. This fossil, first announced in an article in Science magazine in 1994, was a land-dweller which evolutionists, using the same technique of forcing the facts, seek to turn into a whale.
The truth, again, is that there is no evidence that either Pakicetus or Ambulocetus has any relationship to whales. But after these two species, the evolutionary diagram moves on to sea mammals and lists extinct whale species of Archaeoceti such as Protocetus and Rodhocetus. These creatures are indeed extinct sea-dwelling mammals. Yet there are enormous anatomical differences between them and Pakicetus and Ambulocetus. Their fossil remains clearly show that these are not transitional forms linking the species together in any series:

A platypus
Ambulocetus is a four-legged land-dweller. Its backbone ends at the pelvis, to which are connected powerful leg bones. This is the typical anatomy of a land mammal. Whales, however, have no pelvis, and the backbone continues uninterrupted down to the tail. Basilosaurus, believed to have lived up to 10 million years after Ambulocetus, possess the exact same anatomy. In other words, it is a typical whale. No transitional form exists between Ambulocetus, a typical land-dweller, and Basilosaurus, a typical whale.
There are small bones, independent of the verterbrae, in the lower spines of both Basilosaurus and sperm whales. Evolutionists claim that these are “shrunken legs”. In fact though, these bones assisted the adoption of the mating position in Basilosaurus and support the reproductive organs in today’s sperm whales.48 To describe parts of the skeleton that serve an important function as “vestigial” is simply an evolutionist preconception.
In conclusion, the fact remains that marine mammals emerged with all their particular features, with no transitional form between them and land-dwelling mammals. There is no evolutionary chain here at all. Robert Carroll admits as much, albeit reluctantly and using evolutionist terminology: “It is not possible to identify a sequence of mesonychids leading directly to whales.” 49 Despite being an evolutionist, the Russian scientist G. A. Mchedlidze—a well-known expert on the subject of whales—does not agree with Pakicetus, Ambulocetus natans and similar four-legged creatures being described as possible whale ancestors. Rather, he identifies them as a completely isolated group.50
In short, the evolutionists’ scenario—of sea mammals evolving from land-dwellers—is incorrect. Their remaining claim, of evolution amongst sea mammals themselves, also faces a terrible dilemma. Via scientific classification, they attempt to build a family relationship between extinct marine mammals known as Archaeoceti and living whales and dolphins.
Yet experts on the subject think rather differently. The evolutionist paleontologist Barbara J. Stahl writes:
The serpentine form of the body and the peculiar serrated cheek teeth make it plain that these archaeocetes could not possibly have been ancestral to any of the modern whales.51
The evolutionist explanation for the origin of marine mammals is also in a grave predicament presented by findings by molecular biology. The classic evolutionist scenario hypothesizes that the two main whale groups—toothed whales (Odontoceti) and baleen whales (Mysticeti)—evolved from some common ancestor. Michel C. Milinkovitch of the University of Brussels has opposed this view, emphasizing that this assumption is based on anatomical similarity, but invalidated by molecular findings:
While the monophyly of cetaceans is widely accepted, the origin of and evolutionary relationships among the major groups of cetaceans is more problematic since morphological and molecular analyses reach very different conclusions. Indeed, based on the conventional interpretation of the morphological and behavioral data set, the echolocating toothed whales (about 67 species) and the filter-feeding baleen whales (10 species) are considered as two distinct monophyletic groups. . . . On the other hand, phylogenetic analyses of DNA and amino acid sequences contradict this long-accepted taxonomic subdivision. One group of toothed whales, the sperm whales, appears to be more closely related to the morphologically highly divergent baleen whales than to other odontocetes.52
In short, marine mammals contradict every evolutionist scheme into which they are sought to be included.
The Impossibility of any Transition from Land to Sea
As Nature magazine science writer Henry Gee expresses it:
The intervals of time that separate the fossils are so huge that we cannot say anything definite about their possible connection through ancestry and descent.53
There is a generational difference of millions of years separating the fossils claimed to represent the ancestors of marine mammals. Even despite documentary records, it is very difficult to establish the identity of any human’s great-great- great-grandmother, and this sometimes cannot be established at all. For that reason, the claim that fossils representing “transitional forms” are in a relationship of direct linear descent can be only an assumption.
 |
| A Hippopotamus (picture 1)
Imaginary transitional forms with half-hippopotamus,half-whale characteristics (pictures 2-4). There is no trance of such transitional forms in the fossil record. |
Also, it is wrong to try to construct a direct line of decent, based on only a few similarities among species. The striking resemblances between different organisms today were also noticed before Darwin’s time, but were regarded as the product of common design. To suggest that such similarities represent evidence of evolution is not a scientific deduction.
Furthermore, evolutionists need to explain how living species that they claim represent transitional forms could have turned into creatures ideally adapted to the water—and by what mechanisms this came about.
It is not enough merely to claim, “Front legs turned into fins, rear legs disappeared, so did body fur, and fat turned into blubber.” Not a single piece of evidence from present-day creatures can show how front legs could have transformed into fins or how a land-dweller could have adapted perfectly to life in the water, totally changing its body shape and internal bone structure .
No mechanism in nature could carry out the changes that evolutionists maintain took place.
Keeping in mind the infinite adaptations any land-dweller needs to live its life in the sea one can see that even impossible fails to describe such a transition. The absence of just one of these adaptations, claimed to have taken place in the imaginary evolutionary process, will make it impossible for the creature to survive.
The True Origin of Birds
Evolutionists have various scenarios regarding the so-called evolution of birds, though they have no scientific evidence for any of them. According to the most popular one, birds evolved from carnivorous dinosaurs known as Theropods. The ornithologist Storrs L. Olson of the Smithsonian Institution Museum of Natural History refers to this claim, which evolutionists are unable to support with any evidence, as “one of the grander scientific hoaxes of our age.”54
Olson criticizes those who suggest that birds evolved from dinosaurs, although he himself cannot offer an alternative evolutionary explanation for birds’ origin.
In order for a land-dweller to acquire the ability to fly, it would need to undergo a huge number of anatomical and physiological changes. The theory of evolution is unable to explain how these changes might have come about, nor is it able to offer any evidence from the fossil record that such changes took place at all. Therefore, the “birds are feathered dinosaurs” theory is rejected by a number of biologists and paleontologists who nevertheless support the theory of evolution. For example, Alan Feduccia of the University of North Carolina and Larry Dean Martin of the University of Kansas, two of the world’s most prominent ornithologists, don’t believe that birds can have evolved from any known dinosaur group. Feduccia in particular, despite believing in evolution, emphasizes the differences between dinosaurs and birds and produces evidence to show that these are very considerable—for which reason birds could not possibly have evolved from earlier dinosaurs.
 |
| 1:According to the most popular evolutionist claims regarding the origin of birds, they evolved from the theropod dinosaurs illustrated to the side. This scenario is devoid of any evidence.
2: The skeleton of a Herrerasaur, a theropod species. |
To demonstrate why it is that the theory of evolution is a terrible predicament when it comes to the origin of birds, recall some of the differences between birds and reptiles:
1) A bird’s lung possesses an entirely different structure from all other land-dwelling vertebrates, reptiles included. In the bird lungs, unlike in land vertebrates, air travels in just one direction, allowing birds to constantly take in oxygen and expel carbon dioxide. It is impossible for this structure, unique to birds, to have evolved from the land-vertebrate lung, because any transitional reptile-bird would be unable to breathe at all.55
2) In 1992, a comparative study by Alan Feduccia and Julie Nowicki of bird and reptile embryos demonstrated enormous differences between the foot structures of the two groups and that it is impossible to establish any evolutionary link between them.56
 |
| 1: A Complete bird
Transitional forms such as these, with half-dinosaur,half-bird features,never existed.(pictures 2-4)
5: Adrawing of a dinosaur For evolutionists to be able to prove that birds evolved from dinosaurs, they need to find so-called transitional forms of the kind pictured. However, although there are many fossils of dinosaurs and of birds in the fossil record, there is no sign of any imaginary dino-bird. |
3) Recent comparisons of the skulls of the two groups reached the same conclusion. As a result of a 1999 study, Andrzej Elzanowski concluded that “the differences resulted from a phylogenetic reduction rather than individual variation.”57
4) Teeth are one of the factors distinguishing birds from reptiles. It is known that in the past some birds did have teeth in their beaks. This was for long portrayed as evidence of evolution, although it was subsequently realized that bird teeth are quite unique. Feduccia writes on the subject:
Perhaps the most impressive difference between theropods and birds concerns the structure of teeth and the nature of their implantation. . . It is astounding that more attention has not been given to the dramatic differences between bird and theropod teeth (Table 1), especially when one considers that the basis of mammal paleontology involves largely tooth morphology. . . . To be brief, bird teeth (as seen in Archaeopteryx, Hesperornis, Parahesperornis, Ichthyornis, Cathayornis, and all toothed Mesozoic birds) are remarkably similar and are unlike those of theropods . . . There is essentially no shared, derived relationship of any aspect of tooth morphology between birds and theropods, including tooth form, implantation, or replacement.58
5) Birds are warm-blooded, and reptiles cold-blooded. This indicates two completely different metabolisms, and random mutations could not possibly have performed any transition between the two by. (The thesis that dinosaurs were actually warm- blooded was put forward in order to resolve this. Yet a great deal of evidence disproves that thesis, which itself rests on no evidence.)59
6) Reptiles have scales, and birds feathers—two totally different structures. It is impossible for feathers to have evolved from scales.
7) Reptiles have heavy, thick, solid bones. Birds’ bones, on the other hand, are thinner and hollow. Their lighter bones make it possible for birds to fly with ease.
These are just a few of the differences between the two verterbrates. A reptile would have to undergo countless mutations in order to acquire bird characteristics. Just for a reptile’s front legs to develop into wings, for example, it would have to undergo a great many gradual changes. Every mutation affecting the genetic data for the foreleg must make certain minute changes; and with every change, the leg must acquire slightly more wing characteristics. For instance, for feathers to appear on the legs, they must develop gradually: First the stem must emerge, for example, followed by the other components. Toes must disappear a little more with every succeeding generation, and the foot must increasingly come to resemble a wing. And at least some of these very slow, gradual changes must also be observed in the fossil record. The same applies to the creatures’ lungs, changes in the structure of the teeth, and other features.
But mutations lack the ability to perform such wide-ranging, gradual changes. As pointed out earlier, mutations are nearly always harmful. Since they also come about in a random manner, they lack any planning and organization—and of course the conscious intent to transform one organ into another, gradually and accurately at every stage. (For more detailed information, see Harun Yahya, Darwinism Refuted, New Delhi: Goodword Books, November 2002)
Had evolution actually occurred between reptiles and birds, then we should have millions of transitional fossils as evidence. To date, however, not a single half-bird/half-reptile fossil has ever been found. Those that have been discovered belong to either extinct birds or reptiles. The dino-bird stories we so frequently run across in the media are nothing more than sleight of hand, as you shall be seeing in some detail. None of these is the missing link in the so-called evolution of birds.
The Error of Believing Flying Reptiles to be the Ancestors of Birds
Some people, who find the theory of evolution credible only because of their one-sided and second-hand knowledge of it, believe that extinct flying reptiles like pterodactyls are the ancestors of birds. The fact is, flying reptiles have absolutely no connection with birds, and no evolutionist authority maintains that birds evolved from these highly-specialized reptiles.
These flying reptiles are an extinct group known as pterosaurs, whose origin is a major dilemma for the theory of evolution. Like so many other species in the fossil record, they emerge suddenly with all their unique features fully formed. Carroll, one of the world’s foremost authorities on vertebrate paleontology, makes the following confession, in spite of his own evolutionist credentials: “ . . . all the Triassic pterosaurs were highly specialized for flight. . . They provide little evidence of their specific ancestry and no evidence of earlier stages in the origin of flight.”60
The wing structure of flying reptiles is particularly fascinating: There are four fingers on the pterosaur’s wing, just as there are on the front legs of other reptiles. The fourth “little” finger is some 20 times longer than the others, however, and the wing stretches out beneath it. If flying reptiles had evolved from land-dwelling reptiles, then this fourth finger must have grown very gradually, and in stages. Yet not only is there no evidence of this in the fossil record, but neither can any such growth be explained in terms of the natural selection-mutation mechanisms, because the transitional-form stages would make the hand non-functional, without allowing the creature to fly.
 |
| The flying reptiles known as pterosaurs have very different wing and skeletal structures from birds. |
It is a grave error to ascribe an evolutionary relationship between birds and flying reptiles, if only because of their very different wing structures. In the same way that it would be ignorant to claim an evolutionary relationship among flies (insects, bats (mammals) and starlings (birds) on the premise that they all have wings, it is equally erroneous to posit such a relationship between flying reptiles and birds.
Feathered Dinosaur Tales
 |
| Alan Feduccia |
For the last decade, dinosaurs with feathers or “dino-birds” have been one of the most prominent propaganda vehicles in the Darwinist media. Headlines about imaginary dino-birds have followed hot on one another’s heels, and reconstructions and self-confident statements by “experts” have been employed to convince people that half-bird half-reptile creatures once roamed the Earth. But there is not a shred of evidence for this.
Alan Feduccia, of the University of North Carolina Biology Department, is one of the world’s most eminent authorities on the origin of birds, whose name will appear on any list of the world’s five most eminent ornithologists. Dr. Feduccia believes in the theory of evolution, and that birds themselves evolved. But what distinguishes him from the “dino-bird” supporters and certain fanatical evolutionists is that he admits the theory of evolution’s uncertainty on this subject and attaches no credence to the dino-bird scenario, which rests on no foundations at all.
The Auk magazine is published by The American Ornithologists’ Union and represents a floor for the most technical ornithological debates. In its October 2002 edition, Dr. Feduccia’s article “Birds Are Dinosaurs: Simple Answer to a Complex Problem,” provides some important information. He describes in detail how the theory that birds evolved from dinosaurs, advanced by John Ostrom in the 1970s and which has been fiercely defended ever since, lacks any scientific evidence—and why such an evolution is impossible.
Dr. Feduccia is not alone in this. The evolutionist Peter Dodson, a professor of anatomy from University of Pennsylvania, also states that he looks with suspicion on the claim that birds evolved from theropod dinosaurs.61
Feduccia describes an important fact about the “dino-birds” discovered in China: Even if apparently primitive feathers are found on the fossilized reptiles which are portrayed as feathered dinosaurs , it is by no means certain that they are bird feathers. On the contrary, there is considerable evidence that these fossil traces, known as “dino-fuzz,” have nothing to do with bird feathers. As Feduccia writes:
The so-called “hairy devil” pterosaur Sordes (Upper Jurassic lake deposits of Kazakhstan) is preserved in similar lacustrine deposits and preserves structures remarkably similar, if not identical, to dino-fuzz (Wellnhofer 1991). Yet, the finest-grained lithographic limestone, the Archaeopteryx- bearing Solnhofen beds, do not preserve dino-fuzz on the small coelurosaur Compsognathus, a very close ally of the early Cretaceous Chinese Sinosauropteryx, which exhibits a halo of that material.62
Following that analysis, Feduccia avers that some paleontologists are behaving in a prejudiced manner in this view:
. . . shared by many paleontologists: birds are dinosaurs; therefore, any filamentous material preserved in dromaeosaurs must represent protofeathers.63
According to Feduccia, one of the facts invalidating this preconception is traces of dino-fuzz that can have no possible connection with bird feathers:
Most important, “dino-fuzz” is now being discovered in a number of taxa, some unpublished, but particularly in a Chinese pterosaur (Wang et al. 2002) and a therizinosaur, which has teeth like those of prosauropods. Most surprisingly, skin fibers very closely resembling dino-fuzz have been discovered in a Jurassic ichthyosaur and described in detail (Lingham-Soliar 1999, 2001). Some of those branched fibers are exceptionally close in morphology to the so-called branched protofeathers (“Prum Protofeathers”) described by Xu et al.”64
Feduccia recalls that in the past, certain structures were found next to fossils and were originally thought to belong to these fossils—but were later identified as inorganic substances:
One is reminded of the famous fernlike markings on the Solnhofen fossils known as dendrites. Despite their plantlike outlines, these features are now known to be inorganic structures caused by a solution of manganese from within the beds that re-precipitated as oxides along cracks or along [the] bones of fossils.65
Therefore, even if feathered dinosaurs did exist at one time, this still constitutes no evidence that birds evolved from dinosaurs. The “feathers” claimed to have been present on the dinosaurs in question bear no resemblance to birds’ feathers, with their own unique structure, design, and biochemistry. There is definitely no structure resembling bird feathers on these reptiles. According to Professor Alan H. Brush, of the University of Connecticut: “ . . . in development, morphogenesis, gene structure, protein shape and sequence, and filament formation and structure, feathers are different.”66

A drawing of Therizinosaurus
Moreover, since bird feathers are so exceedingly complex, there should be many transitional forms demonstrating the evolution of such a structure. Yet no such transitional form exists. Nature magazine admits this fact:
Feathers are complex structures. Their abrupt appearance in the bird fossil record has been difficult to explain, mainly because no intermediate structures are preserved in the related theropod taxa.67
Therefore, even if a feathered dinosaur were found, it could never be regarded as evidence that birds evolved from dinosaurs, because bird feathers are unique structures, and no evidence suggests that they evolved from anything else.
Also noteworthy is that all the fossils presented as “feathered dinosaurs” were found in China. Why have they all been unearthed in China, and nowhere else in the world? Chinese fossil beds are capable of preserving not just uncertain structures such as dino-fuzz, but bird feathers as well. Feduccia also questions this:
One must explain also why all theropods and other dinosaurs discovered in other deposits where integument is preserved exhibit no dino-fuzz, but true reptilian skin, devoid of any featherlike material, and why typically Chinese dromaeosaurs preserving dino-fuzz do not normally preserve feathers, when a hardened rachis, if present, would be more easily preserved.68
So what are all these so-called “feathered dinosaurs” found in China? What is the true identity of these creatures portrayed as transitional forms between reptiles and birds?
Feduccia explains that some of the animals portrayed as “feathered dinosaurs” are extinct reptiles displaying dino-fuzz, and that others are real birds:
There are clearly two different taphonomic phenomena in the early Cretaceous lacustrine deposits of the Yixian and Jiufotang formations of China, one preserving dino-fuzz filaments, as in the first discovered, so-called “feathered dinosaur” Sinosauropteryx (a compsognathid), and one preserving actual avian feathers, as in the feathered dinosaurs that were featured on the cover of Nature, but which turned out to be secondarily flightless birds.69
In other words, all the fossils presented to the world as feathered dinosaurs or dino-birds belong to either various flightless birds, like today’s ostriches, or else to reptiles possessing the organic material known as dino-fuzz, which has nothing to do with true bird feathers. Not a single fossil represents any transitional form between birds and reptiles.
The Question of Age and the Cladistic Error
In all the evolutionist publications which encourage the dino-bird concept, one important fact is insistently ignored, or even concealed: The ages of the fossils deceptively put forward as dino-birds or feathered dinosaurs go back no further than 130 million years. Yet a true bird, Archaeopteryx, is at least 20 million years older than the creatures evolutionists try to describe as “semi-birds.” Archaeopteryx is known as the world’s oldest true bird, with perfect flight muscles, flight feathers and an authentic bird skeleton, which soared through the skies 150 million years ago. That being so, it is nonsensical to portray creatures which lived long after Archaeopteryx as the ancestors of birds.
However, evolutionists have found a way to defend that nonsensicality: the so-called cladistic method. This term refers to a new method of interpreting fossils, which has been heard frequently in the world of paleontology over the last 20 to 30 years. Proponents of the cladistic method recommend simply ignoring the age of fossils and propose only comparing the characteristic features of fossils we already possess—and constructing evolutionary trees in the light of the similarities that emerge as a result of those comparisons.
 |
| Evolutionists resort to distortions for the sake of constructing so-called evolutionary relationships. For instance, they regard the Velociraptor, which is much younger than Archaeopteryx, as the latter’s ancestor. 1: A drawing of Archaeopteryx
2: AVelociraptor fossil
3: Adrawing of a Velociraptor |
On one evolutionist website, those who support that view explain why it is logical(!) to regard the Velociraptor, a much younger dinosaur than Archaeopteryx, as its ancestor:
Now we may ask, “How can Velociraptor be ancestral to Archaeopteryx if it came after it?” . . . because of the many gaps in the fossil record, fossils don’t always show up “on time.” For example, a recently discovered partial fossil from the Late Cretaceous of Madagascar, Rahonavis, seems to be a cross between birds and something like Velociraptor, but appears 60 mys too late. No-one, however, says its late appearance is evidence against its being a missing link, it may just have lasted a long time [without becoming extinct]. Such examples are called “Ghost Lineages”; we assume these animals existed earlier when we have probable ancient ancestors for them a long way back, and perhaps possible descendants back then too.70
This account is an excellent summary of the cladistic method, revealing what a grave distortion it really is. Evolutionists openly distort results from the fossil record, according to the requirements of their own theories. There can be no meaning in a distortion that assumes that with a 70-million-year-old fossil species actually lived 170 million years ago, and then constructing a family relationship upon that.
Peter Dodson, a professor of anatomy from Pennsylvania University, states that the appearance of so-called dino-birds after the presence of the first true birds represents a serious problem, and that the solution offered by the cladistic method is a forced one:
I continue to find it problematic that the most birdlike maniraptoran theropods are found 25 to 75 million years after the origin of birds? Ghost lineages are frankly a contrived solution, a deus ex machina required by the cladistic method. Of course, it is admitted that late Cretaceous maniraptorans are not the actual ancestors of birds, only “sister taxa.” Are we being asked to believe that a group of highly derived, rapidly evolving maniraptorans in the Jurassic gave rise to birds, as manifested by Archaeopteryx, and then this highly progressive lineage went into a state of evolutionary stasis and persisted unchanged in essential characters for millions of years? 71
The cladistic method is actually a veiled admission of the theory of evolution’s defeat in the face of the fossil record. To summarize:
1) Darwin proposed that once the fossil record was examined in detail, transitional forms would be found to fill the gaps between all the known species. That was his expectation.
2) However, the best efforts of paleontology over the last 150 years have revealed no transitional forms, nor any trace of such creatures. This is a huge upset for his theory.
3) In the same way that no transitional forms have been found, the ages of the creatures which can be announced to be one another’s ancestors solely on the basis of similarities are also contradictory. A creature that appears to be more primitive, according to evolutionists, emerges after one that appears more mature.
This final point forced evolutionists to develop the inconsistent cladistic method. With cladistics, Darwinism clearly demonstrates that it is not a theory based on scientific findings, but rather a dogma that distorts the scientific facts and changes them in accordance with its own assumptions.
The Origin of Birds’ Feathers
Evolutionists maintain that feathers—a feature unique to birds, and an exceedingly complex structure— evolved from reptilian scales. Like the other distinctive features of birds, however, there exist no transitional forms in the fossil record to show how feathers evolved in a gradual process. The fossil record has preserved reptiles’ scales, birds’ feathers, and even mammals’ fur and skin, but no creature has ever been found with part-scale and part-feather structures to prove there was an ongoing, gradual transition to fully-formed feathers.
Some evolutionists maintain that since birds have fragile, hollow bones, they have not left well-preserved fossils behind them. That is most definitely not so case. Birds and their feathers leave behind excellent fossils, especially in regions formerly occupied by ancient lakes, internal bays and shallow seas. As a result, bird fossils are frequently discovered.
In the same way that half-feather-half-scale, or half-skin-half-feather structures have never been found in the fossil record, neither have any with fewer feathers than present-day specimens.72 In an article in American Zoology magazine, Larry Dean Martin, and Stephen. A. Czerkas, director of the Blanding Dinosaur Museum, write, “The oldest known feathers . . . are already modern in form and microscopic detail.” 73
For example, Archaeopteryx is the oldest known bird. Despite having a unique structure different from that of present-day birds, it still has feathers totally resembling those on present-day birds.74
Analysis of Archaeopteryx’s feathers, perfectly preserved from 150 million years ago, concluded that their every detail was exactly the same as present-day bird feathers.75 As far back as 1910, the famous ornithologist and writer on natural history W. P. Pycraft stated that an Archaeopteryx feather was in no respect different to the most developed known present-day feather.76
The rich fossils obtained since then have done nothing to alter his judgment. Besides, we have a lot more information about the skin of the dinosaurs. According to the conclusion reached from an analyses of them, “The skin of a wide variety of dinosaurs is now known and is unlikely to represent a predecessor to a feather-bearing integument.” 77
 |
| Reptile scales (picture 1), of which there are many examples in the fossil record
These imaginary forms with half scale, half-feather features do not exist. (pictures 2-4)
Bird feathers (picture 5) of which there are many examples in the fossil record |
Evolutionists’ claim regarding the way in which bird feathers evolved produced mutually contradictory theories.78 Old textbooks on the subject of evolution refer to imaginary transitional-form bird feathers and suggest that they would soon be found in the fossil record. Yet none of these expected transitional forms have been unearthed so far. Nonetheless, evolutionists still continue to claim that bird feathers evolved from reptiles’ scales. According to these claims, the scales gradually grew longer, grew fronds and slowly assumed a form capable of bearing the bird in such a way as to enable flight.79 However, this is no more than imaginative speculation, on and devoid of any scientific proof.
Actually, since there are so many major differences between bird feathers and reptile scales, there ought to be a great many transitional forms between them. Yet no such fossils appear in the fossil record.80
Bird Feathers in Amber
One of the oldest known bird feathers was found in amber dating back to the Cretaceous Period (144 to 165 million years ago, at the end of the Mesozoic Period). The feather stem and fibers were perfectly preserved, and it was even established which species of bird the feather belonged to. Despite this discovery of a feather dating back 165 million years, there is still no evidence in the fossil record of feathers’ alleged evolution. As one Columbia University biologist put it, “ . . . we lack completely fossils of all intermediate stages between reptilian scales and the most primitive feather.” 81 A great many bird fossils have been found in the fossil record, all with perfect feathers. For that reason, the origin of feathers represents a major question mark for Darwinists.82
 |
| There are many fossils belonging to bird species in the fossil record. |
 |
| Left, a 90- to 95-million-year-old bird feather in amber. Below left, a 120-million-year-old feathered bird fossil, with a bird feather of the same age to its right. |
The True Origin of Man
The origin of mankind is one of the most problematic issues for evolutionists. The human skeleton’s upright posture, the use of our hands, our brain, skull and many more physiological and anatomical features, as well as our human intellect and consciousness—all are very different from those of any other living thing.

As can be seen from this schema, the fossils that evolutionists claim are the ancestors of man have no line of descent relationship showing continuity from the past to the present. Each one emerges as the continuation of a different species.
Evolutionists claim that we humans evolved from an imaginary common ancestor that we share with the apes. But they have yet to explain how the changes necessary for this came about, simply with random mutations, much less show evidence in the fossil record of the stage-by-stage development of every human feature. Actually, they possess not a single fossil to demonstrate the so-called evolution of Man.
The biologist and mathematician Marcel-Paul Schutzenberger summarizes some of the difficulties facing the theory of evolution with regard to the origin of humans:
Gradualists and saltationists alike are completely incapable of giving a convincing explanation of the quasi-simultaneous emergence of a number of biological systems that distinguish human beings from the higher primates: bipedalism, with the concomitant modification of the pelvis, and, without a doubt, the cerebellum, a much more dexterous hand, with fingerprints conferring an especially fine tactile sense; the modifications of the pharynx which permits phonation; the modification of the central nervous system, notably at the level of the temporal lobes, permitting the specific recognition of speech. From the point of view of embryogenesis, these anatomical systems are completely different from one another. Each modification constitutes a gift. . . It is astonishing that these gifts should have developed simultaneously. Some biologists speak of a predisposition of the genome. Can anyone actually recover the predisposition, supposing that it actually existed? Was it present in the first of the fish? The reality is that we are confronted with total conceptual bankruptcy. 83
To conceal their hopeless position regarding the alleged evolution of Man, and also to console themselves, evolutionists set out fossils from various extinct species of apes and human races, in an imaginary order. None of these remains reveals a progression from ape-like creatures to Man. Evolutionists try to give the theory of evolution an alleged scientific appearance and credibility with imaginative models and drawings and biased interpretations of selected fossils.
 |
Henry Gee, editor of Nature, stated in an article in the July 12, 2001, issue that the hominid (human-like) fossils that evolutionists claim to represent the ancestors of modern man, do not follow a progression from the primitive to the more advanced—and that on the contrary, these fossils appear suddenly in the record. The article also explains that transitional forms, awaited for 150 years as proof of the theory of evolution, do not exist, and that different species all emerged suddenly. He uses the following analogy: “Discoveries of fossil hominids are like buses: nothing for a while, then three come along at once.” 84
 |
| Nature, July 12, 2001 |
In his book In Search of Deep Time, Gee states that the so-called plan of human evolution (below) gives no information about forebear-descendant relationships; that there is no missing link; and that human beings are not observed to have followed a gradual development. He also states that the living species in the plan emerged in completely different places.85
Gee again emphasizes the lack of evidence for the so-called evolution of mankind:
Fossil evidence of human evolutionary history is fragmentary and open to various interpretations. Fossil evidence of chimpanzee evolution is absent altogether. 86
He is not alone in making admissions of this kind. In an article in Nature, Professor Bernard Wood of George Washington University writes, that the evolutionary origins of Man are shrouded in darkness:
It is remarkable that the taxonomy and phylogenetic relationships of the earliest known representatives of our own genus, Homo, remain obscure. Advances in techniques for absolute dating and reassessments of the fossils themselves have rendered untenable a simple unilineal model of human evolution, in which Homo habilis succeeded the australopithecines and then evolved via H. erectus into H. sapiens—but no clear alternative consensus has yet emerged. 87
Richard C. Lewontin, professor at Harvard University’s Museum of Comparative Zoology, admits that there is no evidence of so-called human evolution in the fossil record:
When we consider the remote past, before the origin of the actual species Homo sapiens, we are faced with a fragmentary and disconnected fossil record. Despite the excited and optimistic claims that have been made by some paleontologists, no fossil hominid species can be established as our direct ancestor
.. . . The earliest forms that are recognized as being hominid are the famous fossils, associated with primitive stone tools, that were found by Mary and Louis Leakey in the Olduvai Gorge and elsewhere in Africa. These fossil hominids lived more than 1.5 million years ago and had brains half the size of ours. They were certainly not members of our own species, and we have no idea whether they were even in our direct ancestral line or only in a parallel line of descent resembling our direct ancestor. 88
 |
| Evolutionists have put in 150 years of effort in looking for fossils of imaginary transitionalform creatures in order to prove their theory. Yet those 150 years have borne no fruit at all. |
Michael D. Lemonick, the Associate Editor of Time magazine, and a devout evolutionist, describes the despair of his fellows on the subject of human evolution in an article titled “How Man Began”:
Yet despite more than a century of digging, the fossil record remains maddeningly sparse. With so few clues, even a single bone that doesn’t fit into the picture can upset everything. Virtually every major discovery has put deep cracks in the conventional wisdom and forced scientists to concoct new theories, amid furious debate. 89
Significantly, although evolutionists are aware that they have no evidence for evolution at all, still they feel compelled to defend their theory. This goes to show just how fanatical evolutionists are with regard to their theory, and how they can act in violation of both science and reason.
Evolutionists’ Imaginary Ancestors
The idea of human evolution, bereft of any evidence to support it, begins the human family tree with a species of ape known as Australopithecus. It’s claimed that Australopithecus gradually came to adopt an upright posture. Its brain grew larger and after undergoing a number of stages, evolved into modern man, Homo sapiens. Yet the fossil record again fails to back up this scenario. Despite all the claims about transitional forms, an unbreachable barrier remains between human and ape fossils. Moreover, it has also emerged that species formerly depicted as one another’s ancestors actually lived during the same periods.
Australopithecus
Evolutionists refer to mankind’s alleged first ancestors as Australopithecus, meaning “the southern ape.” Although there are various species of Australopithecus, only A. afarensis is actually regarded as a direct ancestor of human beings. (This is the species represented by “Lucy,” discovered in 1974 and announced to the world as proof of evolution.) However, detailed analyses of Australopithecus fossils have revealed that these actually represent an extinct ape species of ape.
| It used to be suggested that lucy was an ancestor of man before it was realized it was actually an etinct species of ape. 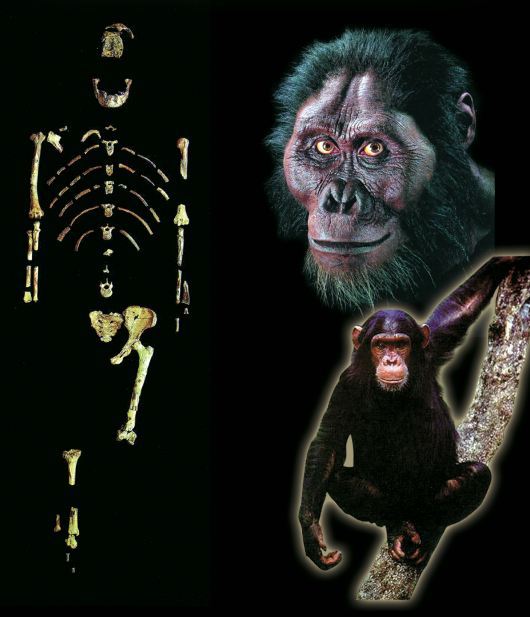 |
| “Lucy,” of the species Australopithecus afarensis, otherwise known as AL 288-1. |
It is thought that the Australopithecines first appeared in Africa some 4 million years ago, and survived until about 1 million years ago. All of these extinct apes resembled present-day monkeys. Their brain volumes are equivalent to or smaller than present-day chimpanzees. They have nodules on the bones of their hands and feet to assist in tree-climbing—again just like modern apes; and their feet are prehensile, to assist in tree-climbing. They are short, about 130 centimeters (51 inches) at most and just like modern apes, male Australopithecines are larger than the females. Many other features—such as hundreds of details in their skulls, eyes close to each other, sharp incisors, the structure of their jaws, long arms and short legs—all show that they are little different from apes living today.
The evolutionist claim is that although Australopithecines possess a completely simian anatomy, they walked upright—unlike modern apes.
In fact, however, many studies on Australopithecus have concluded that the species did not walk upright, and was not bipedal:
1. The world famous anatomist Lord Zuckerman, despite favoring the theory of evolution, concluded that the Australopithecines were merely a species of ape and very definitely did not walk upright.90
2. Charles E. Oxnard, well known for his research on this subject, and his team concluded that Australopithecus’s skeletal structure resembled that of present-day orangutans.91
3. In 1994, Fred Spoor of London’s Global University and his team carried out a wide-ranging study on the Australopithecus skeleton to arrive at a definitive conclusion. The study was performed on an organ in the ear known as the cochlea which determines the body’s position relative to the ground. Spoor reached the conclusion that Australopithecus did not walk in the same manner as human beings.92
4. In 2000, Brian. G. Richmond and David. S. Strait discussed the bones in the Australopithecus forearm in a study published in Nature magazine. Comparative anatomical studies showed that this species had the same forearm anatomy as present-day quadripedal apes.93
Indeed, years before the famous evolutionist Richard Leakey said that the Australopithecine manner of walking resembled that of monkeys:
The Rudolf Australopithecines, in fact, may have been close to the “knuckle-walker’” condition, not unlike the extant African apes 94
Christine Berg, instructor at Washington University School of Medicine, concluded in a 1994 article in the Journal of Human Evolution that the walk and posture of Australopithecus were very different from those of human beings:
The present results lead to the conclusion that the bipedalism of the Australopithecus must have differed from that of Homo. Not only did Australopithecus have less ability to maintain hip and knee extension during the walk, but also probably moved the pelvis and lower limb differently. It seems that the australopithecine walk differed significantly from that of humans, involving a sort of waddling gait, with large rotary movements of the pelvis and shoulders around the vertebral column. Such a walk, likely required a greater energetic cost than does human bipedalism.95
Professor Peter Andrews, of the London Natural History Museum Paleontology Department, states that Australopithecines exhibit more ape-like features and that their foot structure is suited to living in the trees. In his article in Nature magazine, Professor Andrews says:
Developmental patterns were also more ape-like than human. Whether they were phylogenetically hominines or not, it seems to me that ecologically they may still be considered as apes.96
Professor Charles E. Oxnard also accepts that Australopithecines cannot represent a transitional form and are not human-like, but rather constitute a unique group:
In each case although initial studies suggest that the fossils are similar to humans, or at the worst intermediate between humans and African apes, study of the complete fossils clearly differ more from both humans and African apes, than do these two living groups from each other. The australopithecines are unique. 97
 |
| The chimpanzees of Bwindi, which walk on two legs, refute the theory of evolution. |
That Australopithecus cannot be regarded as an ancestor of man is also accepted by the well-known French magazine Science et Vie and other similar publications. The magazine took the subject as the cover story of its May 1999 edition, which dealt with Lucy, regarded as the most important fossil specimen of the species Australopithecus afarensis. Under the headline “Adieu Lucy” (“Farewell, Lucy”), the article stated that the Australopithecus apes were not the ancestors of humans and needed to be removed from the human evolutionary tree.98
One final discovery revealing the invalidity of the thesis that Australopithecus walked on two legs was encountered in the Bwindi jungle in Uganda. The University of Liverpool researcher Robin Crompton discovered that chimpanzees here walk on two legs. In a report in the newspaper The Scotsman under the caption “Chimps on Two Legs Run Through Darwin’s Theory,” Crompton comments that : “This is contrary to the accepted idea that we evolved from chimpanzees which were knuckle-walking—or walking around on all fours.”99
As you see, there’s no reason to regard Australopithecus as the ancestor of Man. Creatures belonging to this species are merely an extinct species of ape.
Homo habilis
That the skeleton and skull of Australopithecus are virtually identical to those of chimpanzees, and the sound evidence disproving the theory that they walked upright left evolutionist paleontologists in a rather difficult position, because in their imaginary scheme, Australopithecus was followed by Homo erectus. As its Latin name suggests, Homo erectus is of the human genus, and its skeleton is completely upright, with a skull volume up to twice as large as that of Australopithecus. Even according to the theory of evolution, it is impossible for there to be any direct transition from a chimpanzee-like ape species such as Australopithecus to H. erectus with its skeletal structure indistinguishable from that of modern humans.
Links, or in other words transitional forms, are therefore necessary. The concept of H. habilis was born of that necessity.
The classification of H. habilis was first suggested in the 1960s by the Leakey family of fossil hunters. According to the Leakeys, this new species possessed the ability to walk upright, a relatively large brain volume, and the ability to use stone and wooden implements—and might therefore be an ancestor of Man.

The Leakey family, known as the “fossil hunters.” Above, Louis and Mary Leakey Left, Richard Leakey
In the mid-1980s, however, newly discovered fossils belonging to the same species altered this view entirely. Based on these fossils, researchers such as Bernard Wood and Loring Brace said that the classification Australopithecus habilis, meaning “tool-using South African ape,” should be employed instead of H. habilis, which means “tool-using human.” This was because H. habilis shared a great many features with the apes of the Australopithecus genus. Just like an Australopithecus, it possessed a long-armed, short-legged and ape-like skeletal structure. Its hands and feet were suited to climbing. Its jaw structure completely resembled those of present-day apes. Its brain volume of 630 cubic centimeters was another indication that this was an ape species. In short, H. habilis, depicted by some as a transitional form, was actually an extinct species of ape—like all other Australopithecines.
Research in subsequent years revealed that H. habilis was in fact a creature no different from Australopithecus. The fossil skeleton and skull referred to as OH62, discovered by Tim White in 1984, showed that like modern apes, this species had a small brain volume, long arms useful for climbing, and short legs.
Detailed analyses by the American anthropologist Holly Smith in 1994 again showed that H. habilis was actually an ape, not a human being. After her analysis of the teeth of the species Australopithecus, H. habilis, H. erectus and H. neandertalensis, Smith said the following:
Restricting analysis of fossils to specimens satisfying these criteria, patterns of dental development of gracile australopithecines and Homo habilis remain classified with African apes. Those of Homo erectus and Neanderthals are classified with humans.100

Australopithecus robustus possesses classical ape features.
That same year, Fred Spoor, Bernard Wood and Frans Zonneveld arrived at the same conclusion by a very different method, based on comparative analyses of the semi-spherical canals in the inner ear of apes and human beings that serve to establish balance. Spoor, Wood and Zonneveld summarized how the first fossils to exhibit human morphology belong to the Homo erectus group, but that Australopithecus—and Paranthropus, known as A. robustus—exhibit classic ape characteristics:
Among the fossil hominids the earliest species to demonstrate the modern human morphology is Homo erectus. In contrast, the semicircular canal dimensions in crania from southern Africa attributed to Australopithecus and Paranthropus resemble those of the extant great apes 101
In their study of the H. habilis fossil Stw 53, Spoor, Wood and Zonneveld found, surprisingly, that “Stw 53 displayed less two-legged behaviour than Australopithecines.” This meant that the H. habilis specimen resembled an ape far more than did Australopithecus. These researchers therefore concluded that Stw 53 represents an unlikely intermediate between the morphologies seen in the Australopithecines and H. erectus.102
In a 1999 article published in Science, Wood and Collard repeated the conclusion arrived at:
We present a revised definition, based on verifiable criteria, for Homo and conclude that two species, Homo habilis and Homo rudolfensis, do not belong in the genus. 103
As a result of their own research, some scientists such as S. Hartwig-Scherer and R. D. Martin stated that H. habilis exhibited far more ape-like features than Australopithecus:
Based on the length of the femur in relation to the length of the humerus (humerofemoral index), the emerging picture is that H. habilis has humerofemoral proportions similar to living African apes...104
Ian Tattersall, in a paper titled “The Many Faces of Homo habilis,” makes this comment:
It is increasingly clear that Homo habilis has become a wastebasket taxon, little more than a convenient recipient for a motley assortment of hominid fossils. 105
To summarize the outcome of all these findings, two important conclusions may be set out:
(1) The fossils known as Homo habilis are actually part of the class Australopithecus, not of the class Homo.
(2) Both H. and Australopithecus walked bent over and had the skeletons of monkeys. They have nothing to do with human beings, and are not transitional forms in the so-called human evolutionary tree.
Homo erectus
Homo erectus means “erect-walking man.” Evolutionists have had to distinguish these humans from earlier ones with the appellation erect. That is because all the H. erectus fossils obtained are upright, unlike Australopithecus or H. habilis: There is no difference between the modern human skeleton and that of H. erectus.
 |
| Homo erectus is not a transitional form, but a member of the human race. A great many people today have skull averages of the same size as that of Homo erectus. This shows that H. erectus is a full human, not a transitional form |
One of the major grounds for evolutionists declaring H. erectus as “primitive” is its small brain volume (900 to 1,100 cubic centimeters) compared to the average for man living today, its narrow forehead and thick eyebrows. However, many people alive in the world today have the same average skull dimension as H. erectus (pygmies, for instance). And various modern races also have narrow foreheads and protruding eyeridges (native Australians, or Aborigines, for example).
It is an established fact that differences in skull volume constitute no difference in terms of intelligence and ability. Intelligence does not vary according to brain size, but according to its internal organization.106
The fossils that introduced H. erectus to the world were those of Peking Man and Java Man, both discovered in Asia. Over time, however, it was realized that these two remains were not reliable. Peking Man consisted only of plaster models, the original having been lost. All that remained of Java Man was a piece of skull and a pelvic bone found several dozen meters away: There was no indication that these had belonged to the same creature. For that reason the H. erectus fossils discovered in Africa gained increasing importance.
Examination of the Turkana Boy, the best-known of these H. erectus fossils discovered in Africa, revealed that there was no difference between H. erectus and present-day Man.
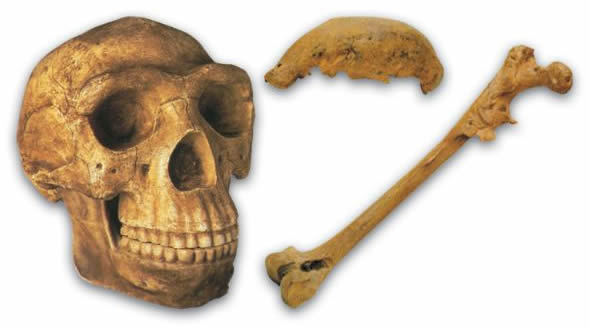 A skull fragment showing that Homo erectus had a large brain. A fossil bone showing that Homo erectus walked erect. PEKING MAN: AN EXAMPLE OF EVOLUTIONIST FRAUD |
Even the evolutionist paleontologist Richard Leakey stated that the difference between H. erectus and present-day Man goes no further than racial difference:
One would also see differences: in the shape of the skull, in the degree of protrusion of the face, the robustness of the brows and so on. These differences are probably no more pronounced than we see today between the separate geographical races of modern humans. Such biological variation arises when populations are geographically separated from each other for significant lengths of time.107
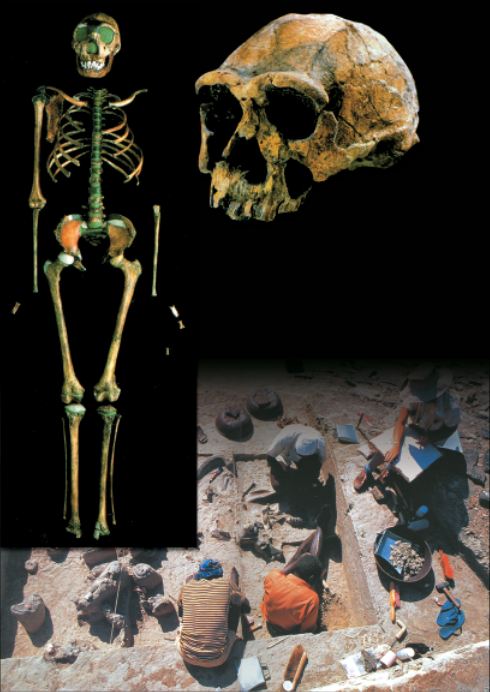 |
| THE “TURKANA BOY,” ADVANCED AS A TRANSITIONAL FORM FOSSIL, IS ACTUALLY NO DIFFERENT FROM PRESENT-DAY MAN... The best known of the Homo erectus specimens found in Africa is the fossil known as the Turkana Boy. The fossil has been established as having belonged to a 12-year-old child. The erect structure of the skeleton is no different from that of man living today. |
Professor William Laughlin of University of Connecticut spent years performing anatomical research on the Inuit (formerly known as Eskimos) and the people of the Aleutian Islands, and saw that these people bore a surprising resemblance to H. erectus. Laughlin concluded that all these “species” were in reality different races of H. sapiens—or present-day Man:
When we consider the vast differences that exist between remote groups such as Eskimos and Bushmen, who are known to belong within the single species of Homo sapiens, it seems justifiable to conclude that Sinanthropus belongs within this same diverse species.108
Increasingly, scientific magazines are referring to H. erectus as being an artificial classification, and to the fossils included within the category H. erectus as insufficiently different from H. sapiens to be considered a separate species. American Scientist summarizes the debate on the issue and the outcome of a conference held in 2000:
. . . most of the participants at the Senckenberg conference got drawn into a flaming debate over the taxonomic status of Homo erectus, started by Milford Wolpoff of the University of Michigan, Alan Thorne of the University of Canberra and their colleagues. They argued forcefully that Homo erectus had no validity as a species and should be eliminated altogether. All members of the genus Homo, from about 2 million years ago to the present, were one highly variable, widely spread species, Homo sapiens, with no natural breaks or subdivisions. The subject of the conference, Homo erectus, didn’t exist. 109
Scientists who support this thesis reached the conclusion that H. erectus is not a different species, but a race within Homo sapiens.
There is a huge gulf between H. erectus, a human race, and the apes that precede it in the “human evolution” scenario (Australopithecus, H. habilis and H. rudolfensis). To put it another way, the first humans identified in the fossil record appeared suddenly and simultaneously, with no evolutionary process.
Homo sapiens archaic, Homo heilderbergensis and Cro-Magnon
In the imaginary evolutionary tree, H. sapiens archaic represents the stage before modern day Man. In fact, there is little to say about this subspecies from the point of view of evolution, since they are distinguished from modern day Man by only very minor differences. Indeed, some researchers point to the native people of Australia, saying that representatives of this race (H. sapiens archaic) are still alive today. Like that race, native Australians have thick bones behind the eyebrows, an undershot jaw and a slightly smaller brain volume. Furthermore, a number of very serious findings show that in the very recent past, these people also lived in some villages in Hungary and Italy.
The class called Homo heilderbergensis in evolutionist literature is actually the same thing as H. sapiens archaic. The reason for this use of two different names to describe the same human race is differences of opinion amongst evolutionists. All the fossils classed under H. heilderbergensis show that people closely resembling modern Europeans, anatomically speaking, lived in England and Spain 500,000 and even 780,000 years ago.
 |
| The 780,000-year-old human fossils discovered in the Gran Dolina Cave in northern Spain were classified as Homo heilderbergensis. |
Cro-Magnon man is estimated to have lived up to some 30,000 years ago. This race possessed a dome-shaped head and a broad forehead. Their skull volume of 1600 cubic centimeters is greater than the modern day average. There are large eyebrow protrusions in the skull and at the back, a bony protrusion typical of Neanderthal Man and H. erectus.
Cro-Magnons are regarded as a European race, yet the structure and volume of their skulls more closely resemble those of certain races living today in Africa and tropical climates. Based on this similarity, Cro-Magnon man is believed to be an ancient race originating in Africa. A number of other paleoanthropological findings show that the Cro-Magnon man and Neanderthal races interbred, forming the rootstock of some modern day races.
In conclusion, these people are neither “primitive species” nor transitional forms, but different races of humans who lived in the past and either fused and were assimilated with other races, or else became extinct.
 |
| FOSSILS FROM DIFFERENT HUMAN RACES ARE DEPICTED BY EVOLUTIONISTS AS HALF-APE, HALF-HUMAN. The fossil record contains fossils belonging to different human races or species of ape. However, there is no trace of the half-ape, halfhuman creatures so beloved of evolutionists. picture 1:A Cro-Magnon skull
picture 2:A Neanderthal skull
picture 3:People of different human races |
Human beings are always human beings in the fossil record, and apes are always apes,
As we have shown so far, information obtained from the fossil record shows that the human “evolution” scenario has no scientific foundations. What is found in the fossil record is fossil remains of either human beings, or else of apes. There is no trace of the transitional forms hoped for by evolutionists. Indeed, no mechanism exists in nature that might bring about such an evolution. The theory of evolution is unable even to explain how a single protein molecule could have arisen by chance, and it is definitely out of the question for it to account for the evolution, as a result of chance mutations, of human beings, with their complex bodies, ability to think, rejoice, decide, comprehend, take pleasure from art and beauty, compose music, write books, and all their other characteristics.
In short, there is no evidence that human beings came into existence through evolution. Such a gradual change is in any case impossible. The fact that evolutionists are unwilling to accept that fact alters nothing. The Creator of Man is not blind chance, but Almighty God, the Great and Mighty, Lord of the Worlds.

Say: "Have ye seen (these) 'Partners' of yours whom ye call upon besides Allah. Show Me what it is they have created in the (wide) earth. Or have they a share in the heavens? Or have We given them a Book from which they (can derive) clear (evidence)?- Nay, the wrong-doers promise each other nothing but delusions. (Surat al Fatir, 40)

