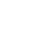
H
H
Haeckel, Ernst
 |
| Ernst Haeckel |
The famous evolutionist biologist Ernst Haeckel was a close friend and supporter of Darwin. To support the theory of evolution, he put forward the theory of recapitulation, which maintained that the embryos of different life forms resembled one another in their initial stages. It was later realized that in putting forward that claim, Haeckel had produced forged drawings. (See Embryological evolution.)
While perpetrating such scientific frauds, Haeckel also engaged in propaganda in favor of eugenics. He was the first to adopt and disseminate the idea of eugenics in Germany. (See Eugenic slaughter.) He recommended that deformed newborns babies should be killed without delay and that the evolution of society would thus be accelerated. He went even further, maintaining that lepers, patients with cancer and the mentally ill should be ruthlessly done away with, lest such people prove to be a burden on society and slow its evolution.
George Stein summarized Haeckel’s blind devotion to the theory of evolution:
. . . [Haeckel] argued that Darwin was correct . . . humankind had unquestionably evolved from the animal kingdom. . . humankind’s social and political existence is governed by the laws of evolution, natural selection, and biology, as clearly shown by Darwin. To argue otherwise was backward superstition. 197
Hallucigenia
This is one of the life forms that emerged suddenly in the Cambrian Period in its perfect form. (See Cambrian Period.) This Cambrian fossil has sharp spines to protect it against attack—and evolutionists are unable to explain how this creature came to have such excellent protection at a time when there were no predators around. The absence of predators makes it impossible to account for this in terms of natural selection.
 |
| This Hallucigenia fossil, one of the life forms that suddenly emerged in the Cambrian Period, has spines for protection |
Heterotrophic view, the
The heterotrophic view is one of the most researched theses regarding the emergence of the first life. According to this view, a consuming life form absorbs from its outside environment the organic molecules it needs for the formation of structures and to meet its energy requirements. This theory maintains that the first life form fed on organic compounds that formed spontaneously within a highly complex framework. It had no need for a gene system to enable it to synthesize the simple organic molecules it absorbed from the environment. In other words, this hypothetical first living thing was able to maintain its vital functions as a simple-structured feeder in a complex environment.
According to this view, chemical evolution took place before life formed. As the result of the lengthy evolution of inanimate substances, heterotrophic life forms emerged. Again according to this view, there was no free oxygen in the primeval atmosphere. The gasses assumed to have existed then—ammonia (NH3), methane (CH4), hydrogen (H2) and water vapor (H20)—underwent chemical reactions with high-energy ultraviolet rays and gave rise to more complex compounds. At the end of these reactions, the substances that emerged by chance first combined in tiny droplets of water and were gradually transported to the seas and oceans where, it’s assumed, they gave rise to simple organic compounds.
 |
| The wings of the modern bird Opisthocomus hoatzin have claw-like talons, just like Archaeopteryx |
All the research conducted to confirm this hypothesis has ended in failure. This has not even been possible in controlled laboratory experiments, let alone as the work of chance. (See Fox Experiment, the and Miller Experiment, the.)
Hoatzin Bird, the
The points on which evolutionist base their portrayal of Archaeopteryx as a transitional form are its skeleton, which resembles that of dinosaurs, the claws on its wings, and the teeth in its mouth. (See Archaeopteryx.) They therefore claim that Archaeopteryx was a transitional form that still had many reptilian features, but had newly acquired some bird-like ones.
However, the “reptilian” features in question do not actually make Archaeopteryx a reptile at all. The claims put forward pointing to its claws are particularly invalid, because there are birds with clawed wings alive today. Just like Archaeopteryx, the Australian Hoatzin has clawed wings.198 Again like Archaeopteryx, it flies with a small breastbone. However, for that reason alone, evolutionists claim that Archaeopteryx was unable to fly, or could not fly very well. This demonstrates that such features as claws, teeth and skeletal structure like those in Archaeopteryx make it a unique species of bird, not a reptile.
However, all kinds of biased interpretations can be made from the evolutionist perspective. Were a fossil Hoatzin discovered today in the appropriate geological strata, very likely it would be proposed as a transitional form, in the same way as Archaeopteryx was. But the fact that this creature is still alive and manifestly a bird does not let evolutionists make any such claim.
Homo Antecessor
The most astonishing fact to tear up the imaginary tree of life by its very roots is the way that Homo sapiens dates back to unexpectedly early times. Paleontological findings show that human beings identical to the H. sapiens were living nearly a million years ago.
The findings on this subject 199 were rejected by some evolutionist paleoanthropologists, because of their damaging implications for the evolutionary family tree. One fossil discovered in Atapuerca in Spain in 1995 revealed in a most striking manner that H. sapiens was far older than had been thought. (See Atapuerca.) This fossil indicated that the history of H. sapiens needed to be put back to at least 800,000 years ago. However, once they had got over their initial shock, evolutionists decided that the fossil belonged to a different species, because—according to the evolutionary family tree—H. sapiens could not have been alive 800,000 years ago. They therefore came up with an imaginary species known as Homo antecessor, to which they ascribed the Atapuerca skull.
Homo Erectus
Evolutionists regard the classification Homo erectus, meaning “upright-walking human,” as the most primitive species on the fictitious human family tree. They have had to separate these humans from other, earlier classes by means of the title upright, because all the H. erectus fossils we have are erect in a manner not seen in specimens of Australopithecus or Homo habilis. There is no difference between H. erectus skeletons and those of modern human beings.
Evolutionists’ most important grounds for regarding H. erectus as “primitive” are the fact that its brain volume (900 to 1100 cubic centimeters) is smaller than the modern human average, and also its thick protruding eyebrow ridges. The fact is, however, that a great many human beings today have a brain size identical to that of H. erectus (pygmies, for example), and eyebrow protrusions can also be seen in various contemporary human races, such as native Australians. It is a known that there is no correlation between brain size and intelligence and ability. Intelligence varies not according to brain size, but according to its internal organization. 200
| Homo Erectus’ Maritime Culture |
 |
| According to “Ancient Mariners: Early humans were much smarter than we suspected” an article published in New Scientist magazine on 14 March 1998, the human beings whom evolutionists refer to as Homo erectus were building boats some 700,000 years ago. It is of course out of the question to regard people possessed of boatbuilding knowledge as primitive. |
The fossils that introduced H. erectus to the world were Peking Man and Java Man fossils, both discovered in Asia. However, it was gradually realized that these two remains were not reliable. (See Java Man, Peking Man.) For that reason, more and more importance began to be attached to the H. erectus fossils discovered in Africa. (Also, some evolutionists included some of the fossils described as H. erectus in a second class, Homo ergaster. by. The matter is still a subject of debate.)
The best-known of the H. erectus specimens discovered in Africa is Nariokotome homo erectus or the so-called Turkana Boy. The fossil’s upright skeleton is identical to that of modern man.201 Therefore, H. erectus is a human race that is still in existence today. (See Turkana Boy, the.)
Professor William Laughlin of the University of Connecticut carried out lengthy anatomical research into Inuit and the inhabitants of the Aleut Islands and noted that these people bore an astonishingly close resemblance to H. erectus. Laughlin’s conclusion was that all these races are actually different races all belonging to H. sapiens, ¸or today’s man:
When we consider the vast differences that exist between remote groups such as Eskimos and Bushmen, who are known to belong to the single species of Homo sapiens, it seems justifiable to conclude that Sinanthropus [an erectus specimen] belongs within this same diverse species [H. sapiens]. 202
| Homo Erectus’ Maritime Culture |
 |
| According to “Ancient Mariners: Early humans were much smarter than we suspected” an article published in New Scientist magazine on 14 March 1998, the human beings whom evolutionists refer to as Homo erectus were building boats some 700,000 years ago. It is of course out of the question to regard people possessed of boat-building knowledge as primitive. |
There is an enormous gulf between Homo erectus, a human race, and the apes that precede it (Australopithecus, Homo habilis, H. rudolfensis) in the scenario of human evolution. In other words, the first humans to appear in the fossil records emerged suddenly, all at the same time, and in the absence of any process of evolution. There could be no clearer indication that they were created.
However, acceptance of this fact would constitute a violation of evolutionists’ dogmatic philosophies and ideologies. Therefore, they seek to depict H. erectus, a human race, as a semi-ape. That is why they insist on giving H. erectus an ape-like appearance in the reconstructions they produce. (For detailed information, see The Evolution Deceit by Harun Yahya.)
Homo ergaster
Some of the fossils described as Homo erectus (“upright-walking human”) are classified as Homo ergaster by certain evolutionists. There is no agreement among evolutionists regarding this second classification. (See Homo erectus.)
Homo habilis
The fact that Australopithecus is nearly identical to chimpanzees in terms of their skull and skeletal structures, plus the emergence of concrete evidence that demolished the claim that these creatures walked upright, left evolutionist paleoanthropologists in a rather difficult position. Because Australopithecus is followed by Homo erectus in the imaginary course of evolution.
As can be seen from the use of the term Homo in its Latin name, Homo habilis is a human group and has a completely upright skeleton. Its skull volume is up to double that of Australopithecus. According to the imaginary family tree, H. erectus with a skeleton identical to that of modern man, coming immediately after Australopithecus, a species of ape similar to chimpanzees, cannot be explained in terms of the theory of evolution. Therefore, links or transitional forms are necessary. It is this difficulty that gave rise to the concept of Homo habilis.
 |
In the 1960s, the Leakeys, the fossil hunter family, proposed the classification Homo habilis. According to the Leakeys, this new species classified as H. habilis possessed the ability to walk upright, had a relatively large brain, and was able to use tools made of stone and wood. It may therefore have been an ancestor of modern man.
New fossils belonging to the same species discovered after the mid-1980s totally altered that view. Researchers such as Bernard Wood and Loring Brace said that the term Australopithecus habilis or “tool-using South African ape” should be employed instead of Homo habilis, which means “tool-using human,” because H. habilis shared a great many characteristics with the apes known as Australopithecus.
Just like Australopithecus, it had a long-armed, short-legged and ape-like skeletal structure. Its fingers and toes were suited to climbing. Its jaw structure completely resembles that of modern apes. Its 500-cubic-centimeter brain size was the best indication that it was in fact an ape. In short, H. habilis, portrayed as a separate species by some evolutionists, was actually an ape just like all other Australopithecus.
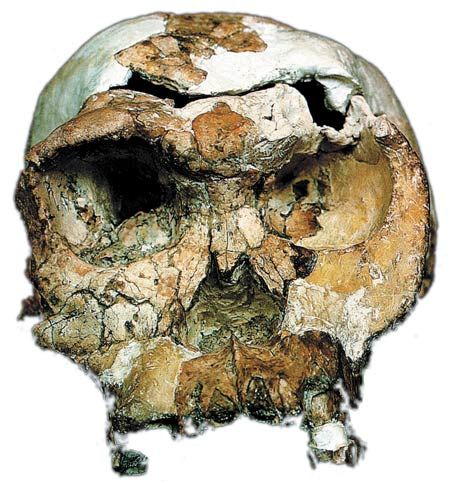
 |
| Analyses of the teeth of the species Australopithecus and Homo habilis (at side) shows that these belong to the same category as African apes. |
Detailed analyses conducted by the American anthropologist Holly Smith in 1994 again showed that H. habilis was not homo at all, but rather a monkey. Smith said this about these analyses of the teeth of Australopithecus, Homo habilis, H. erectus and H. neandertalensis:
Restricting analysis of fossils to specimens satisfying these criteria, patterns of dental development of gracile australopithecines and Homo habilis remain classified with African apes. Those of Homo erectus and Neanderthals are classified with humans. 203
That same year, three experts in anatomy—Fred Spoor, Bernard Wood, and Frans Zooneveld—arrived at the same conclusion using another method. This was based on a comparative analysis of the semi-spherical canals that serve to establish balance in the inner ears of human beings and monkeys. The inner ear canals of all the Australopithecus and H. habilis specimens that Spoor, Wood, and Zonneveld examined by were identical to those of modern apes. That of H. erectus, on the other hand, was the same as that of modern human beings.
This finding led to two conclusions:
1. The fossils known as Homo habilis actually belong to the classification Australopithecus, not to Homo, or modern man.
2. Both H. habilis and Australopithecus walked with a stoop, and had monkey-like skeletons. They have nothing to do with human beings.204
Homo heidelbergensis
The classification referred to as H. heidelbergensis in evolutionist literature is actually the same as Homo sapiens archaic. The reason why these two separate names are used to describe the same human race is the differences of opinion among evolutionists. All the fossils included under the classification Homo heidelbergensis show that human beings very anatomically similar to modern Europeans were living in Britain and Spain 500.000 and even 740,000 years ago.
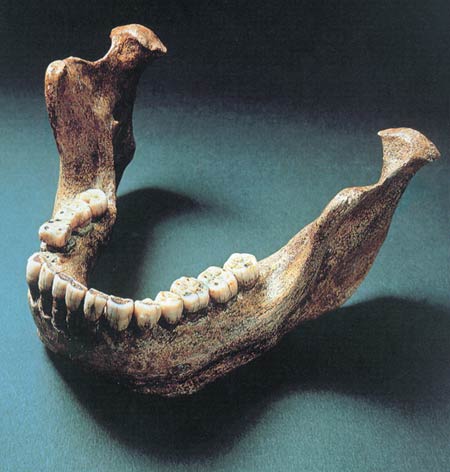 |
| Homo heilderbergensis fossils show that human beings bearing a close resemblance to modern Europeans were living in Britain and Spain some 740,000 years ago. |
Homo rudolfensis
This is the Latin name given to a few fossil fragments discovered in 1972. Since these were unearthed near the River Rudolf in Kenya, the species they were assumed to represent was given the name Homo rudolfensis. The majority of paleoanthropologists, however, regard these fossils not as a new species but as H. habilis—in other words, a species of monkey.
Richard Leakey, who discovered the fossils, presented the skull—which he estimated to be 2.8 million years old and which was given the official designation of KNM-ER 1470, as the greatest discovery in the history of anthropology—and thus aroused an immense reaction. According to Leakey, this creature, with a small skull volume like that of Australopithecus and with a human-type face, was the missing link between Australopithecus and man.
However, it was later realized that the human facial features of KNM-ER 1470, which had appeared on the covers of various scientific journals, were errors, made perhaps even deliberately in assembling the skull fragments. Professor Tim Bromage, who conducted research into the anatomy of the human face, summarized his findings produced with the aid of computer simulations in 1992:
When it [KNM-ER 1470] was first reconstructed, the face was fitted to the cranium in an almost vertical position, much like the flat faces of modern humans. But recent studies of anatomical relationships show that in life the face must have jutted out considerably, creating an ape-like aspect, rather like the faces of Australopithecus. 205
On the same subject, the evolutionist paleoanthropologist J. E. Cronin says this:
... its relatively robustly constructed face, flattish naso-alveolar clivus, (recalling australopithecine dished faces), low maximum cranial width (on the temporals), strong canine juga and large molars (as indicated by remaining roots) are all relatively primitive traits which ally the specimen with members of the taxon A. africanus..... KNM-ER 1470, like other early Homo specimens, shows many morphological characteristics in common with gracile australopithecines that are not shared with later specimens of the genus Homo. 206
C. Loring Brace from Michigan University arrived at the following conclusion regarding the KNM-ER 1470 skull, based on analyses of its jaw and teeth structure:
. . . from the size of the palate and the expansion of the area allotted to molar roots, it would appear that ER 1470 retained a fully Australopithecus -sized face and dentition. 207
John Hopkins University Professor of paleoanthropology Alan Walker, who has examined the KNM-ER 1470 skull at least as much as Leakey, maintains that like Homo habilis or H. rudolfensis, the creature should not be classified as Homo but should be included in the classification Australopithecus.208
Classifications such as Homo habilis or H. rudolfensis, which evolutionists depict as transitional forms between Australopithecus and Homo erectus, are completely fictitious. Most present-day researchers accept the fact that these are members of the series Australopithecus. All their anatomical characteristics point to the fact that they were monkeys.
Homo sapiens
The history of Homo sapiens, which represents modern man on the imaginary evolutionary family tree, goes back much further than evolutionists expected. Paleontological findings indicate that human beings identical to us were alive nearly a million years ago.
One of the discoveries in this regard is a fossil found in the Atapuerca region of Spain. The fact that this fossil has the same characteristics to those of modern man rocked evolutionist beliefs regarding the evolution of man. Because according to the evolutionary family tree, no Homo sapiens should have lived 800,000 years ago.
Indeed, many findings showed that H. sapiens goes back even further than 800,000 years. One of these was the discoveries made in Olduvai Gorge by Louis Leakey in the early 1970s. Leakey determined in the Bed II stratum that Australopithecus, Homo habilis and H. erectus had all lived there together and at the same time.
However, the really interesting thing was a structure—the remains of a stone hut—found by Leakey in that same stratum (Bed II). Its most interesting aspect was that such a structure, which is still in use in some African regions today, could have been made only by Homo sapiens! According to Leakey’s findings, Australopithecus, Homo habilis, H. erectus and modern man must all have been living together around 1.7 million years ago. 209This fact of course invalidates the theory of evolution that maintains that modern human beings evolved from the monkeys described as Australopithecus.
Moreover, there are findings of traces of modern human beings that go back even further than 1.7 million years. The most important of these is the footprints found in the Laetoli region. (See Laetoli footprints, the.) These prints, identical to those of modern human beings, have been calculated to date back some 3.6 million years.
These footprints discovered by Mary Leakey were later examined by such well-known paleoanthropologists as Don Johanson and Tim White.
Examinations of the morphological structure of the footprints again showed that they had to be regarded as belonging to a human and, what is more, to modern man, Homo sapiens. Russell Tuttle investigated the prints and later wrote:
A small barefoot Homo sapiens could have made them. . . In all discernible morphological features, the feet of the individuals that made the trails are indistinguishable from those of modern humans. 210
Unbiased investigations described the owners of these two sets of footprints: There was a total of 20 fossilized footprints belonging to a human being aged around 10, and 27 fossilized footprints belonging to someone slightly younger. They were very definitely normal human beings just like ourselves.
The fact that evolutionists persist in their theory that clearly conflicts with the scientific findings, and the way that they distort or ignore every discovery that works against it, clearly reveals that theory’s unscientific nature.
Homo sapiens archaic
Homo sapiens archaic represents the rung before modern man on the illusory evolutionary ladder. In fact, evolutionists have nothing to say about these people because they are distinguished from modern man only by very minute differences. Some researchers even suggest that representatives of this race are still alive today, citing native Australians as an example. Just like this race, native Australians have heavy eyebrow ridges, a retracted jaw structure and a rather smaller brain volume. And native Australians are a normal human race. (See Aborigines.)
Homology (Common origins)
In biology, structural similarities among different living species are referred to as homologous. Evolutionists attempt to use these similarities as evidence for evolution. Pointing to homologous organs in different life forms, they maintain that these species are descended from a common forebear. (See Homologous organs.) Yet in order for evolutionist claims regarding homologous organs to be taken seriously, these organs would have to be coded by similar DNA codes. Yet these homologous organs are generally determined by different genetic (DNA) codes.
In addition, similar genetic codes in different life forms also correspond to very different organs! In his book Evolution: A Theory in Crisis, the Australian professor of biochemistry Michael Denton describes the predicament represented by the evolutionist interpretation of homology:
Homologous structures are often specified by non-homologous genetic systems, and the concept of homology can seldom be extended back into embryology. 211
In order for that same claim to be taken seriously, the embryological development process of these similar structures—in other words, the phases of development in the embryo in the mother’s womb—have to be parallel to one another. Yet the embryological stages for similar organs are different in all living things.
 |
| At the molecular level, no organism is the ancestor of any other, nor more primitive or advanced than any other. |
Genetic and embryological research has shown that the concept of homology, which Darwin took as proof that living things are descended from a common ancestor, does not in fact provide any backing for such a definition. Thus it is that science has revealed the unrealistic nature of yet another Darwinist thesis.
The evolutionist claim regarding homology is not only invalid at the level of organs, but also at the molecular level. (See Molecular homology thesis, the.) There are enormous molecular differences between living things that outwardly appear very similar and closely related to one another. Professor Michael Denton comments:
Each class at a molecular level is unique, isolated and unlinked by intermediates. Thus, molecules, like fossils, have failed to provide the elusive intermediates so long sought by evolutionary biology . . . At a molecular level, no organism is “ancestral” or “primitive” or “advanced” compared with its relatives . . . . There is little doubt that if this molecular evidence had been available a century ago. . . the idea of organic evolution might never have been accepted. 212
Homologous organs
Anyone examining the different living species on Earth will observe that there are certain similar organs and characteristics among species. This phenomenon has attracted the notice of biologists ever since the 18th century, but the first to link it to the theory of evolution was Darwin, who maintained that there was an evolutionary link between living things with similar (i.e. homologous) organs, and that such organs were a legacy from their common ancestor.
| Two Unrelated Mammals with Giant Teeth |
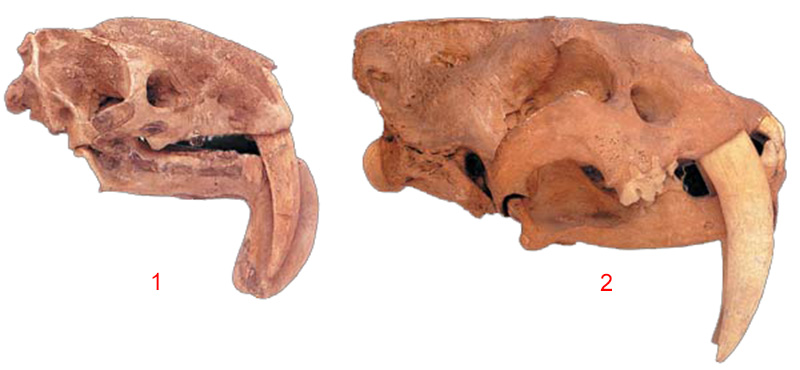 |
| The existence of extraordinarily similar species, one a placental and the other a pouched mammal, deals a severe blow to the claim of homology. For example, Smilodon (right) and Thylacosmilus (left) both have very large front teeth. The fact that the skull and tooth structures of these two life forms, between which no evolutionary relationship can be established, are exceedingly similar refutes |
Accordingly, since pigeons and eagles have wings, that means that pigeons, eagles and other such winged birds all evolved from a common forebear.
Homology is a superficial hypothesis put forward solely on the basis of external appearances. The hypothesis has not been confirmed by any concrete finding since Darwin’s time. In particular, no trace has ever been found of the imaginary common ancestors of life forms with homologous structures as proposed by evolutionists. And there are additional hurdles:
1. The existence of homologous organs in life forms belonging to totally different classes, among which evolutionists also cannot establish any evolutionary link
2. The fact that such homologous organs have very different genetic codes, and
3. The fact that the stages of the embryological development of these organs all show that homology represents no basis for evolution.
Among the examples of species between which evolutionists cannot establish any evolutionary link but which possess homologous structures are those with wings. The bat—a mammal—has wings, and so do birds, and there were once species of dinosaurs that also had wings. However, not even evolutionists can construct any evolutionary relationship among these three different classes.
Another striking example in this context is the astonishing similarity and structural resemblance between the eyes of different living things. For example, the octopus and man are two very different life forms—mollusk and mammal—between which no evolutionary link can be construed. In terms of their structure and function, however, their eyes are actually very similar. Not even evolutionists will claim that human beings and octopi had a common ancestor with a similar eye. These and countless other similar examples make it clear that there is no scientific basis to the evolutionist claim that homologous organs prove that living things are descended from a common evolutionary ancestor. In fact, these organs represent a major impasse for them.
“Hopeful Monster” theory, the
The “Hopeful Monster” theory claims that one day, a reptile laid an egg and that quite by chance, a creature with brown fur hatched out of it. According to evolutionists, when this mammal grew up, it found a mate that had also suddenly emerged from a reptile egg—and a new species resulted.
The reaction from scientists with any common sense ran along the lines of “Is this a scientific account, or a Greek myth, or a Hans Christian Anderson fairy tale?” Yet for some reason, a number of scientists still imagine that it represents a solution to an evolutionary problem. What it actually represents, however, is total despair. One paleontologist, Otto Schindewolf, proposed that all major evolutionary transformations must have occurred in single large steps, like a reptile laying an egg from which a bird hatched.213 As you see, some evolutionists believe that a perfectly formed but totally different species can hatch out of any viable egg!
Of course, the sudden emergence of different living groups in the fossil record showed that living species did come into being with no evolutionary process behind them.214 Naturally, this was a source of major concern for evolutionists.
The “Hopeful Monster” theory was put forward in the 1930s by the European paleontologist Otto Schindewolf, who proposed that living things evolved not with the accumulation of small mutations over time, as neo-Darwinism maintained, but by sudden and very large ones. (See Macro-Mutation deception, the.) In citing examples for his theory, Schindewolf claimed that the first bird had emerged from a reptile egg by way of a gross mutation—by some enormous, random change in its genetic structure. 215
According to his theory, some land animals may have turned into giant whales through sudden and wide-ranging changes. Schindewolf’s fantastical theory was adopted and supported in the 1940s by the Berkeley University geneticist Richard Goldschmidt, although it was so inconsistent that it was swiftly abandoned.
But due to the lack of any transitional forms in the fossil record, the Harvard University paleontologists Stephen Jay Gould and Niles Eldredge were again obliged to resuscitate the idea to account for that situation. Gould’s famous article “Return of the Hopeful Monsters” was an expression of this obligatory about-face.216
Although they did not repeat Schindewolf’s theory to the letter, Gould and Eldredge sought to come up with a mechanism for sudden evolutionary leaps in order to endow the theory with a scientific gloss. (See Punctuated evolution myth, the.) In the years that followed, Gould and Eldredge’s theory was adopted by some other paleontologists, who duly fleshed out its bones. In fact, however, that the theory of punctuated evolution was based on even greater inconsistencies and contradictions than the neo-Darwinist theory of evolution.
Huxley, Julian
 |
| Julian Huxley |
In his 1958 book Religion without Revelation, the zoologist Julian Huxley, one of the architects of neo-Darwinism, described it not as a scientific theory, but as a ideological dogma. (See Neo-Darwinism.)
Hypothesis
A hypothesis is a temporary solution or explanation proposed for a problem raised by various facts or phenomena. In order for a good hypothesis to be confirmed, it needs to be open to experiment and observation, besides conforming to the available facts. At the same time, it must be open to new facts and estimates, and partial changes must be made if required. 217
Scientists first generalize from the observations they carry out or describe a possible cause-and-effect relationship between a chain of events in order to give a temporary explanation for those observations. The first step towards research is taken by means of a hypothesis. Assumptions made in forming that hypothesis must be capable of being tested through controlled experiments. If a hypothesis cannot be based on experimental testing that may confirm it, it remains mere speculation.218
A theory is constructed with a hypothesis, supported by a great many observations and experiments (See Theory.) and includes hypothesis and observation in various different disciplines. For example, the theory of evolution includes hypotheses and observations from paleontology, anatomy, physiology, biochemistry, genetics and other sciences. When a scientist makes an observation that is not compatible with the hypothesis, he must conclude that either the hypothesis or his experiment is flawed. If the observation is correct, he rejects the hypothesis, or else refashions it.
The most important thing in science is for every new observation to conform to the hypothesis.
When it comes to the theory of evolution, however, it appears not to be supported by any hypothesis in any branch of science. Nonetheless, this is wholly ignored for the sake of keeping the theory viable. (See, Evolution theory, the.)
Footnotes
197.George Stein, “Biological science and the roots of Nazism,” American Scientist, Vol. 76(1), 1988, p. 54. 
198.J. Lear Grimmer, National Geographic, August 1962, p. 391.
199.L.S. B. Leakey, The Origin of Homo sapiens, ed. F. Borde, Paris: UNESCO, 1972, pp. 25- 29; L.S.B. Leakey, By the Evidence, New York: Harcourt Brace Jovanovich, 1974.
200.Marvin Lubenow, Bones of Contention, Grand Rapids: Baker, 1992, p. 83.
201.Boyce Rensberger, The Washington Post, November 19, 1984.
202.Marvin Lubenow, Bones of Contention, p. 136.
203.Holly Smith, American Journal of Physical Anthropology, Vol. 94, 1994, pp. 307-325.
204.Fred Spoor, Bernard Wood, Frans Zonneveld, “Implication of Early Hominid Labryntine Morphology for Evolution of Human Bipedal Locomotion,” Nature, Vol. 369, June 23, 1994, pp. 645-648.
205.Tim Bromage, New Scientist, Vol. 133, 1992, pp. 38-41.
206.J. E. Cronin, N. T. Boaz, C. B. Stringer, Y. Rak, "Tempo and Mode in Hominid Evolution," Nature, vol. 292, 1981, pp. 117.
207.C. L. Brace, H. Nelson, N. Korn, M. L. Brace, Atlas of Human Evolution, 2nd Edition, New York: Rinehart and Wilson, 1979.
208.Alan Walker, Scientific American, Vol. 239 (2), 1978, p. 54.
209.A. J. Kelso, Physical Anthropology, 1st Edition, 1970, p. 221; M. D. Leakey, Olduvai Gorge, Vol. 3, Cambridge: Cambridge University Press, 1971, p. 272.
210.I. Anderson, “Who made the Laetoli Footprints?” New Scientist, Vol. 98, 1983, p. 373.
211.Michael Denton, Evolution: A Theory in Crisis, p. 145.
212.Ibid., pp. 290-291
213.Dr. David N. Menton, “The Hopeful Monsters of Evolution,” http://www.gennet. org/facts/metro12.html.
214.Ibid.
215.Stephen M. Stanley, Macroevolution: Pattern and Process, San Francisco: W. H. Freeman and Co., 1979, pp. 35, 159.
216.S. J. Gould, "Return of the Hopeful Monster," The Panda's Thumb, New York: W. W. Norton Co., , 1980, pp. 186-193.
217.Musa Özet, Osman Arpacı, Ali Uslu, Biyoloji 1 (“Biology 1”) Istanbul: Sürat Publishing, 1998, p. 7.
218.Prof. Dr. Ali Demirsoy, Yaşamın Temel Kuralları, Genel Biyoloji/Genel Zooloji, Vol. I, Part I, Ankara, 1993, pp. 12-13.