The NAS’s Error in Portraying Common Structures As Evidence of Evolution
Another phenomenon that the National Academy of Sciences represents as proof of the theory of evolution is homology. Homologies are common structures possessed by different living things. The NAS has taken the similarities in the skeletons of human beings and such animals as mice and bats as an example and proposed that "they are best explained by common descent." (Science and Creationism, p. 14) The NAS repeats the claims made and examples cited in Darwin's The Origin of Species, but entirely ignores the discoveries made in the fields of anatomy and biology since Darwin's day, thus demonstrating that it has remained at the scientific level of 150 years ago.
Before moving on to the NAS's unscientific claims, let us first have a quick look at the concept of homology.
Darwin's Homology Error
In the chapter of The Origin of Species entitled "Mutual Affinities of Organic Beings: Morphology, Embryology, Rudimentary Organs," Darwin spoke of similar structures in species and suggested that this could only be accounted for by his theory of development from a common ancestor.
Richard Milton's book Shattering the Myths of Darwinism
Although Darwin and the evolutionists who came after him maintained that the only explanation for common structures between living things is evolution from a common ancestor, most scientists before Darwin agreed that common structures were the work of a common design.
Darwinists of the past and present regard evolution from a common ancestor as the cause of homology; at the same time, they also portray homology as the strongest evidence for descent from a common ancestor. However, advances in such fields as anatomy, biochemistry, and microbiology over the last 50 years have shown that homology does not constitute evidence for the theory of evolution, and that descent from a common ancestor is not the cause of homology. In his book Shattering the Myths of Darwinism, the well-known science writer Richard Milton states that homology had been one of evolutionists' most important pieces of evidence, but that as science advanced over the course of the twentieth century, homology came to represent one of the most important difficulties facing Darwinism:
In the past hundred years, biology has undergone successive revolutions—in embryology, in microbiology, in molecular biology, and in genetics, revolutions which have laid open on the laboratory bench the most minute detail of how plants and animals are constructed. If the Darwinian interpretation of homology is correct, then you would expect to find at the microscopic level the same homologies that are found at the macroscopic level. In fact that is not what has been found.1
This chapter examines the NAS's claims on the subject of homology and why it represents such a major problem for the theory of evolution. The questions that will be dealt with in more detail in the pages which follow are, in summary:
1. Evolutionists both portray homology as proof of descent from a common ancestor and describe it as descent from a common ancestor. This is a tautology—in other words, circular reasoning—and scientifically speaking proves nothing at all.
2. There are also common structures among living things for which evolutionists do not claim an evolutionary relationship—like those between marsupial and placental mammals, for example. This means that the reason for common structures is not a common ancestor.
3. Contrary to what the NAS would have one believe, the similar structures in many living things are not caused by similar genes. This shows that they do not have a common evolutionary origin.
4. In living things with homologous organs, the developmental stages of these organs are very different. This again shows that these organs do not come from a common ancestor.
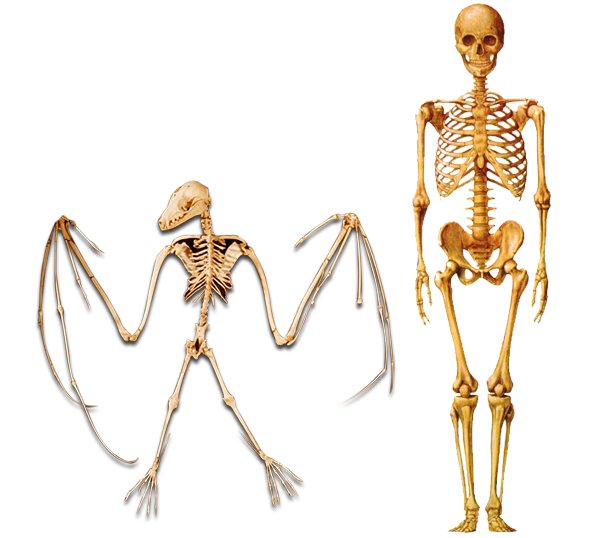
The NAS suggests that the skeletons of such living things as human beings and bats are similar and that this is evidence of evolution. However, this is an error without scientific foundation.
Homology as Evidence for Evolution: An Example of Circular Reasoning
The NAS makes the same mistake as Darwin and suggests that a common ancestor is the best explanation of the common structures among living things. In suggesting that descent from a common ancestor is a certain fact, the NAS is making an assumption, based on a preconception;then it turns around and says that the only explanation for common structures is a common ancestor.
Another error which Darwinists make with regard to homologous organs is hidden in the circular reasoning they employ. According to Darwin and his loyal followers in the NAS, common structures are both the result and proof of the theory of evolution. This deficient logic basically says that the theory of descent from a common ancestor proves homology, which in turn proves descent from a common ancestor.
This is like saying first: "All red convertibles resemble each other; this is proof of the existence of a factory that produces all red convertibles," and then going on to say, "The best explanation of the fact that all red convertibles resemble each other is that they were all made in the same factory." There is no proof of anything here, merely an unproven hypothesis and phenomena interpreted in the light of that hypothesis.
This circular reasoning seen in many of the theory of evolution's claims, as with natural selection, is criticized by many biologists and philosophers. One of these is Ronald Brady, a professor of philosophy from New Jersey's Ramapo College, who wrote the following in 1985:
By making our explanation into the definition of the condition to be explained, we express not scientific hypothesis but belief. We are so convinced that our explanation is true that we no longer see any need to distinguish it from the situation we were trying to explain. Dogmatic endeavors of this kind must eventually leave the realm of science.2

In the same way that it is illogical to claim that all convertible cars were produced in the same factory, it is also illogical to regard living things with similar organs and structures as evidence of descent from a common ancestor.
Why Common Structures Do Not Prove A Common Ancestor
The evolutionists' homology thesis rests on the logic of building an evolutionary relationship between living things with similar structures. The fact is, however, that species between which no evolutionary relationship can be constructed also possess very similar organs. The wing is one example of this. Bats (which are mammals), birds, and flying insects all have wings. Furthermore, in the past there were also winged, flying reptiles. However, it is impossible to construct any evolutionary link or relationship between these four different groups.
Another striking example of this phenomenon is the surprising structural and other similarities in the eyes of living things. The squid and man, for instance, are very different living things, between which no evolutionary link can possibly be established. However, in terms of structure and function their eyes are very similar. Yet not even evolutionists can claim that man and the squid have a common ancestor with similar eyes.
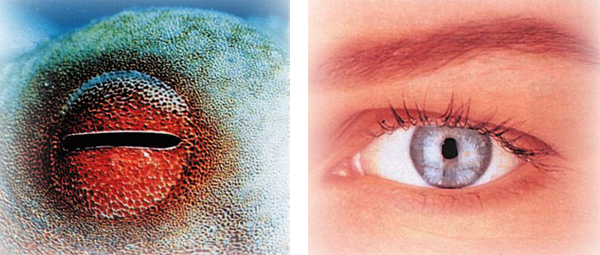
The squid and man are two living things between which it is impossible to build any evolutionary link. Despite this, however, their eyes closely resemble one another in terms of structure and function. This invalidates the evolutionist claim that "common structures reveal a common ancestor."
In the face of this, evolutionists say that these are not "homologous" organs (i.e., descended from a common ancestor), but "analogous" (very similar despite not having an evolutionary connection).
For instance, the human eye and the squid eye are analogous organs in their view. However, the question of whether a given organ will be deemed a homology or an analogy is answered totally in the light of the theory of evolution's preconceptions. This shows that there is nothing scientific about the evolutionist claims based on similarity. Evolutionists interpret discoveries unreservedly according to their dogma, and refuse to behave objectively.
Yet the interpretation they come up with is a most inconsistent one. This is because organs they are forced to regard as "analogous" sometimes resemble each other so closely, despite their extraordinarily complex structures, that it is quite inconsistent to maintain that this similarity came about as the result of chance mutations. If, as evolutionists claim, the squid eye emerged by chance, how is it that exactly the same coincidences took place in the vertebrate eye? The well-known evolutionist Frank Salisbury, who pondered this question long and hard, writes:
Even something as complex as the eye has appeared several times; for example, in the squid, the vertebrates, and the arthropods. It's bad enough accounting for the origin of such things once, but the thought of producing them several times according to the modern synthetic theory makes my head swim.3
According to evolutionist theory, wings emerged four times, totally independently of one another: in insects, flying reptiles, birds and flying mammals (bats). This four-fold emergence of the wing, which cannot be explained by natural selection and mutation mechanisms, as well as the structural similarities between the various kinds of wings, represents a major difficulty for evolutionists.

The existence of similar wings among living things that evolutionists themselves admit are not evolutionarily related represents another dilemma for them.
One of the most concrete examples that totally undermines the evolutionist thesis in this area can be seen in mammals. Modern biologists are agreed that all mammals are divided into three categories: placentals, marsupials, and monotremes. Evolutionists assume that this division goes back to the very beginning and that both categories have a completely independent evolutionary history. How interesting it is, therefore, that there are almost identical "pairs" in placentals and marsupials. Wolves, cats, squirrels, ant-eaters, moles, and mice all have their marsupial counterparts with closely similar morphologies.4 In other words, according to the theory of evolution, completely independent mutations must have twice chanced to produce these living things in exactly the same way! This represents a terrible dilemma for evolutionists.
One of the fascinating similarities between the placental and marsupial categories is that betweenthe North American Wolf and the Tasmanian Wolf. The former is placental and the latter marsupial. Evolutionary biologists believe that these two different species have a totally different evolutionary history.5 (It is assumed that relations between marsupials and placentals have been severed since the Australian continent and the islands around it split away from Gondwanaland, the supercontinent that is supposed to be the originator of Africa, Antarctica, Australia, and South America.) The interesting thing, however, is that the skeletons of the North American and Tasmanian wolves are almost identical. Their skulls resemble each other particularly closely.
Similarities such as these, which evolutionary biologists are unable to accept as "homologous," show that similar organs are no evidence of descent from a common ancestor. Even more interesting, the situation is the exact reverse in some other living things. In other words, there are living things that possess totally different organs despite being regarded as close relatives by evolutionists.
For instance, the great majority of species in the crustacean class have a refracting eye structure. Only two species, the lobster and the shrimp, have a totally different reflecting, mirrored eye structure.

A Tasmanian wolf A North American wolf
1) The skull of a Tasmanian wolf 2) The skull of a North American wolf
Common Structures Controlled by Different Genes
According to the claims made by the NAS and evolutionists on the subject of homology, similar structures and functions in living things must again be controlled by similar genes. As we know, the theory of evolution suggests that living things developed by way of small, random changes in their genes—that is, by mutations. The genetic structures of living things regarded as close evolutionary relatives should therefore also be similar. In particular, similar organs should be controlled by very similar genetic structures. Yet, the fact is that genetic research has revealed results completely at variance with this evolutionist thesis.
Michael Denton's book Evolution: A Theory in Crisis
Similar organs are generally formed by very different genetic (DNA) codes. In addition, similar genetic codes in the DNA of different living things relate to very different organs. In the chapter headed "The Failure of Homology" in his book Evolution: A Theory in Crisis, the Australian biologist Michael Denton states that homology does not offer proof of evolution. Denton states that in order for homology to represent evidence of evolution it needs to show that similar organs are controlled by similar genes and also that similar organs undergo a similar embryological process. However, Denton goes on to say that this is not the case and that homology represents a failure for the theory of evolution:
The validity of the evolutionary interpretation of homology would have been greatly strengthened if embryological and genetic research could have shown that homologous structures were specified by homologous genes and followed patterns of embryological development… But it has become clear that the principle cannot be extended in this way. Homologous structures are often specified by non-homologous genetic systems and the concept of homology can seldom be extended back into embryology. 6
In the same book, Denton summarizes his conclusions:
The evolutionary basis of homology is perhaps even more severely damaged by the discovery that apparently homologous structures are specified by quite different genes in different species.7
In a 1997 article, Richard Milton describes how molecular biology has shattered evolutionists' hopes regarding homology:
It isn't only embryology that experienced such disappointments. In the 1950s, when molecular biologists began to decipher the genetic code, there was a single glittering prize enticing them. When they found the codes for making proteins out of amino acids, they naturally assumed that they were on the brink of discovering at the molecular level the same homologies that had been observed at the macroscopic level in comparative anatomy.
If the bones of the human arm could be traced to the wing of the bat and hoof of the horse, then the miraculous new science of molecular biology would trace the homologies in DNA codes that expressed these physical characteristics… Yet when biologists did begin to acquire an understanding of the molecular mechanism of genetics, they found that apparently homologous structures in different species are specified by quite different genes.8
In fact, this was well known long before. In 1971, the famous evolutionist Gavin de Beer wrote:
It is now clear that the pride with which it was assumed that the inheritance of homologous structures from a common ancestor explained homology was misplaced.9
Work on unraveling the genetic code revealed that anatomical and morphological similarities between living things were non-existent at the genetic level.
De Beer had proven with a number of examples that homologous structures could emerge from different genes. One of these was the segments in insects' bodies. The development of these body parts is controlled by different genes in the fruit fly, the locust, and the wasp. Since it is accepted that the bodies of all insects are homologous, this shows that there is no need for homologous features to be controlled by equivalent genes. Another example is the gene known as "sex-lethal," necessary for sex determination in the fruit fly. This gene is not required for the emergence of males and females in other insects. 10
Another matter which shows that similar organs are not controlled by similar genes is that a gene is generally responsible for more than one feature of a living thing. The gene which determines the color of a rat's fur also determines its dimensions. The gene which determines the color of the fruit fly Drosophila's eyes also controls the female's sex organ. Nearly all the genes in higher organisms have more than one function. The evolutionist biologist Ernst Mayr admits that there are very few or even no genes controlling only one feature.11
Denton cites examples of a pleiotropic gene (having more than one effect) from chickens. The effects of a rather harmful mutation in a single gene may include irregular wing development, lack of toes, sparse feathers, and lung and air-sac deficiencies. The importance of this is that while some affected features such as wings and feathers are specific to birds, others, such as the lung, apply to many other vertebrates, including human beings. Denton stresses "that nonhomologous genes are involved to some extent in the specifications of homologous structures."12
The exact opposite of this—that is, the emergence of non-homologous structures from identical genes—is also frequently encountered. For instance, the gene known as Distal-less is related to the development of limbs in mice, moths, spiny worms, velvet worms, and sea urchins, and yet the appendages of these creatures are all very different. In other words, they are not homologous.
Biologists studying these similarities announced in 1997 that it was astonishing that the appendages of these animals should be so different, and that their anatomies and "evolutionary pasts" must be totally different. In 1999, Professor Gregory Wray of Duke University's Zoology Department found "surprising" the association between the gene Distal-less and "what are superficially similar, but non-homologous structures." Wray's conclusions were;
This association between a regulatory gene and several non-homologous structures seems to be the rule rather than the exception.13
In his book Homology: An Unsolved Problem, published as far back as 1971, the evolutionary biologist De Beer put forward a wide-ranging analysis of the subject and summarized why homology represented a serious difficulty for the theory of evolution:
What mechanism can it be that results in the production of homologous organs, the same "patterns," in spite of their not being controlled by the same genes? I asked this question in 1938, and it has not been answered.14
The question put by De Beer in 1938, to which he was unable to find an answer in 1971, is still unanswered today.
Common Structures With Different Developmental Patterns

Another piece of evidence that undermines the claims of homology is the question of embryological development. Despite the fact that the NAS authors insist in the chapter named "Similarities During Development" that there are similarities between living things during their development and that this is proof of descent from a common ancestor (Science and Creationism, p. 17), this claim does not reflect the true facts.
In order for the evolutionist thesis regarding homology to be taken seriously, the developmental processes of homologous structures—in other words the stages of embryological development in the egg and the mother's womb—need to be parallel. The fact is, however, that these embryological process for homologous organs are very different in every living thing. No matter how much evolutionists choose to ignore it, this truth has been known to scientists since the nineteenth century. For example, the American embryologist E.B. Wilson wrote in 1894,
It is a familiar fact that parts which closely agree in the adult, and are undoubtedly homologous, often differ widely in larval or embryonic origin either in mode of formation or in position, or in both.15
Sixty years after Wilson, De Beer repeated the fact and stated,
The fact is that correspondence between homologous structures cannot be pressed back to similarity of position of the cells in the embryo, or of the parts of the egg out of which the structures are ultimately composed, or of developmental mechanisms by which they are formed.16
This still applies today. The contemporary biologist Pere Alberch makes the following analysis:
[It is] the rule rather than the exception that homologous structures form from distinctly dissimilar initial states. 17
The evolutionary developmental biologist Rudolf Raff studied two species of sea urchin which had reached almost identical forms by way of very different paths, and expressed the same difficulty in 1999:
Homologous features in two related organisms should arise by similar developmental processes . . . [but] features that we regard as homologous from morphological and phylogenetic criteria can arise in different ways in development.18
The incompatibility between the developmental pathway of homologous organs also applies to some vertebrate limbs. Salamanders are one example of this. The development of the digits of many vertebrate limbs is from the back to the front, i.e., from the tail to the head. This is the case with frogs, for instance. Yet, the manner of development of salamanders—which, like frogs, are amphibians—is very different. In salamanders, the development of digits is in the exact opposite direction, from the head to the tail.
Another instance of homologous organs that do not pass through the same embryological stages relates to the way in which these organs generally start to develop in different regions of the embryo. Research has shown that similar organs in different animals begin to be formed by different groups of cells within the embryo. The development of the alimentary canal is one example of this; this fundamental structure emerges in very different ways in a great many different creatures. For example, the alimentary canal in sharks forms from the roof of the embryonic gut cavity. In the lamprey, an eel-like fish, however, it forms from the floor of the gut. In frogs, it begins to form from the floor and roof of the embryo, while in birds and reptiles it starts in the lower layer of the embryonic disc or blastoderm. 19
Many frogs begin life as swimming tadpoles, and turn into frogs during the last stage of metamorphosis.
Darwin's classic example of homology was the forelimbs of vertebrates. This, too, represents a problem for the theory of evolution. This is because the forelimbs emerge in different body segments in different species. In the newt, for example, the forelimbs emerge from segments 2, 3, 4, and 5 of the trunk; in lizards, from segments 6, 7, 8, and 9; and in human beings, from segments 13, 14, 15, 16, 17, and 18.20As the molecular biologist Michael Denton has stated, it could be concluded from this fact that forelimbs are not homologous.21
According to Denton,
The development of the vertebrate kidney appears to provide another challenge to the assumption that homologous organs are generated from homologous embryonic tissue. In fish and amphibia the kidney is derived directly from an embryonic organ known as the mesonephros, while in reptiles and mammals the mesonephros degenerates towards the end of embryonic life and plays no role in the formation of the adult kidney, which is formed instead from a discrete spherical mass of mesodermal tissue, the metanephros, which develops quite independently from the mesonephros...22
The emergence of similar structures as the result of totally dissimilar processes is frequently encountered, especially in the later stages of development.
Many animal species undergo a process known as "indirect development" on the path to adulthood; in other words, they have a larval stage. For example, many frogs start life as swimming tadpoles and turn into four-footed animals at the last stage of metamorphosis. There are also other frog species which bypass the tadpole stage and develop directly. However, most adults from these directly-developing species are almost indistinguishable from other frogs that go through the tadpole phase. 23
In short, embryological and genetic research shows that the concept of homology, which Darwin put forward as proof that living things had developed from a common ancestor, actually represents no such proof at all. A close study of homology demonstrates that it is a clearly inconsistent evolutionist error.
After citing examples from embryology of the dilemma that homology poses for the theory of evolution, Richard Milton says,
Many other comparable examples can be given from embryology: in almost every case they have been put into a file drawer labeled "unresolved problems of homology" and largely forgotten about.24
The way in which the NAS ignores facts known to and accepted by all scientific circles, and tries to portray discredited evidence for the theory of evolution as the definitive truth, is really astonishing.
The Nas's Errors Regarding Reptiles and Mammals
At the end of the chapter on "Common Structures" in the NAS book Science and Creationism, it is stated that "The mammalian ear and jaw are instances in which paleontology and comparative anatomy combine to show common ancestry through transitional stages." (Science and Creationism, p. 15). In essence, the NAS's claim in this chapter is that while the mammalian lower jaw consists of a single bone, the reptile jaw consists of three. Evolutionists maintain that that the greater number of bones in reptiles' ears are homologous to the bones in those of mammals, and put this forward as evidence that mammals evolved from reptiles. One proof of this transition, according to evolutionists and the NAS, is the so-called transitional form Therapsida, a group of mammal-like reptiles with a double jaw joint.
The above account is a classic evolutionist claim. If evolutionists see the slightest similarity between two species, they are not slow to interpret that similarity in terms of evolution. However, in doing so they ignore all the facts that make their interpretation impossible.
The alleged evolution of reptiles into mammals is a matter that contains several major difficulties for evolutionists. The fact that two mammal bones resemble certain bones in reptiles does not resolve the issue. Many questions remain unanswered. For example, how did jaw bones "migrate" to such an irreducibly complex organ as the ear, as a result of mutations? How did these mutations manage to shrink the two jaw bones, cause them to have them the ideal shape and dimensions, and form muscles around them? How did random changes build a perfect balance in the middle ear? And, finally, how did the animal manage to hear and eat while all this was going on? All of these questions remain unanswered. Evolutionists are unable to answer them, because any one of them is sufficient to undermine the myth of the evolution of reptiles into mammals.
Fossils of creatures belonging to the order Therapsida cannot substantiate the evolutionists' claims. First and foremost, therapsid fossils do not appear in the fossil record in the sequence expected by Darwinism. For the evolutionists' claims to be true, therapsid fossils would need to appear in order in the strata from the most reptile-like to the most mammal-like, with respect to jaw features. Yet, this order does not appear in the fossil record.
In his book Darwin On Trial, the famous critic of Darwinism Phillip Johnson makes this comment on the subject:
An artificial line of descent [between reptiles and mammals] can be constructed, but only by arbitrarily mixing specimens from different subgroups, and by arranging them out of their actual chronological sequence.25
In addition, the ear and jaw bones are the only common feature between therapsids and mammals. When the enormous differences between the therapsid and mammalian reproductive system and other organs are examined, it will be seen how far the question of the supposed evolution of reptiles into mammals is from being answered. Taking it still further, matters become even more complicated. For example, one may ask how mammals—a group that includes a great many different categories, such as primates, horses, bats, whales, polar bears, squirrels, and ruminants—evolved from reptiles by means of random mutations and natural selection. (For more detailed information on evolutionists' dilemmas with regard to the origin of mammals, see Harun Yahya, Darwinism Refuted, Goodword Books, 2003.)