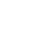
Die wirkliche naturgeschichte - 2 (vögel und säugetiere)
Die Wirkliche Naturgeschichte – Ii (Vögel Und Säugetiere)
Es gibt tausende Vogelarten auf der Erde. Jede einzelne von ihnen besitzt besondere Eigenschaften. Zum Beispiel haben Falken extrem scharfe Augen, weite Schwingen und scharfe Krallen, während Kolibris mit ihren langen Schnäbeln den Nektar aus Blumen saugen.
Andere ziehen über weite Entfernungen zu besonderen Orten der Welt. Doch die wichtigste, die Vögel von anderen Tieren unterscheidende Eigenschaft, ist ihre Fähigkeit zu fliegen.
Wie kamen die Vögel ins Dasein? Die Evolutionstheorie versucht diese Frage mit einem langen Szenario zu beantworten. Nach dieser Geschichte sind Reptilien die Vorfahren der Vögel. 150 bis 200 Millionen Jahre zuvor entwickelten sich Vögel aus Reptilien. Die ersten Vögel konnten nicht sehr gut fliegen. Doch während des Evolutionsprozesses ersetzen Federn die dicke Haut der Urvögel, die ursprünglich mit Schuppen bedeckt war. Die Vorderbeine bedeckten sich ebenfalls mit Federn und veränderten sich zu Flügeln. Als Ergebnis allmählicher Evolution passten sich einige Reptilien an die Anforderungen des Fluges an und wurden zu den heutigen Vögeln.
Dieses Szenario wird in evolutionistischen Quellen als feststehende Tatsache präsentiert. Doch eine tiefergehende Untersuchung der Details und der wissenschaftlichen Daten zeigt, dass das Szenario mehr auf Wunschdenken und Vorstellungskraft, als auf Realität beruht.
Die evolutionistische Auffassung über den Vogelflug
Wie Reptilien als an Land lebende Kreaturen jemals ans Fliegen kamen, ist ein Thema, dass für erhebliche Spekulationen unter Evolutionisten gesorgt hat. Es gibt zwei Haupttheorien. Die erste besagt, die Vorfahren der Vögel seien von den Bäumen auf den Boden hinabgestiegen. Diese Vorfahren sollen Reptilien gewesen sein, die in den Baumkronen lebten und allmählich Flügel entwickelten, weil sie ständig von Ast zu Ast sprangen. Diese Theorie ist bekannt als die Baumtheorie. Die andere, die Lauftheorie legt nahe, dass Vögel sich vom Boden aus in die Luft entwickelten.
Doch beruhen beide Theorien auf spekulativen Interpretationen, und es gibt keinerlei Beweis, der eine von ihnen unterstützen würde. Evolutionisten haben eine einfache Lösung für dieses Problem: Sie stellen sich einfach vor, die Beweise würden existieren. Professor John Ostrom, Chef der geologischen Abteilung an der Yale Universität, der die Baumtheorie vorgeschlagen hat, erklärt diese Auffassung:
Für den Vorläufer der Vögel existiert kein Beweis in Gestalt eines Fossils. Es ist ein rein hypothetischer Vor-Vogel, doch einer, der existiert haben muss.106
Doch diese Übergangsform, von der die Baumtheorie annimmt, sie "muss gelebt haben", ist nie gefunden worden. Die Lauftheorie ist noch problematischer. Ihre Grundnahme ist, die Vorderbeine einiger Reptilien hätten sich bei dem wiederholten Versuch, Insekten zu fangen, zu Flügeln entwickelt. Doch keine Erklärung wird abgegeben darüber, wie denn der Flügel, ein hochkompliziertes Organ, sich durch dieses Umherschlagen mit den Vorderbeinen gebildet haben soll.

IMAGINÄRE THEORIEN, IMAGINÄRE KREATUREN
Die erste von den Evolutionisten vorgeschlagene Theorie zur Erklärung des Ursprungs des Fliegens besagte, Reptilien hätten Flügel entwickelt in dem Versuch, Fliegen zu fangen (oben); die zweite Theorie war, dass sie sich zu Vögeln entwickelten, weil sie von Ast zu Ast sprangen (links). Es gibt jedoch weder Fossilien von Tieren, die allmählich Flügel entwickelt hätten, noch irgendeine Entdeckung, die zeigen würde, dass so etwas auch nur im Bereich des Möglichen liegt
Ein großes Problem für die Evolutionstheorie ist die nicht weiter reduzierbare Komplexität von Flügeln. Nur ein perfekt designter Flügel funktioniert, ein halb entwickelter Flügel kann nicht funktionieren. Aus diesem Grund ergibt der einzigartige, zentrale Mechanismus, nämlich der Mechanismus der "graduellen Entwicklung", der von der Evolutionstheorie behauptet wird, in diesem Zusammenhang keinen Sinn. So ist Robert Carroll gezwungen, einzugestehen:
Es ist schwierig, die Anfänge der Evolution von Federn als Elemente des Flugapparates zu erklären, weil nicht erkennbar ist, wie sie funktioniert haben könnten, bevor sie Dimensionen erreicht hatten, die wir beim Archäopteryx sehen."107
Er argumentiert dann weiter, Federn könnten sich zu Isolationszwecken entwickelt haben, doch dies erklärt nicht ihr komplexes Design, das speziell für das Fliegen perfekt ausgelegt ist.
Wesentlich ist, dass Flügel fest mit der Brust verbunden sein müssen, und so aufgebaut sind, dass sie den Vogel nach oben ziehen können, dass er sich in alle Richtungen bewegen kann und dass er sich in der Luft halten kann. Um dies leisten zu können, müssen sie eine leichte, flexible und ausgewogene Struktur haben. Hier befindet sich die Evolutionstheorie wieder in Verlegenheit. Sie kann die Frage nicht beantworten, wie dieses fehlerlose Design als ein Ergebnis sich angesammelt habender Zufallsmutationen zustandegekommen sein soll. Genauso fehlt die Erklärung, wie die Vorderbeine eines Reptils sich durch einen genetischen Defekt (Mutation) in einen perfekten Flügel verwandelt haben sollen. Ein halb ausgebildeter Flügel kann seinem Besitzer nicht zur Flugfähigkeit verhelfen. Selbst wenn wir also annehmen, dass Mutationen zu einer leichten Veränderung des Vorderbeins geführt haben, ist es doch völlig unvernünftig anzunehmen, dass weitere Mutationen zufällig zur Entwicklung eines voll ausgebildeten Flügels beitrugen. Denn eine Mutation der Vorderbeine wird nicht automatisch einen Flügel ergeben. Sie wird zunächst einmal lediglich dazu führen, dass das betroffene Tier seine Vorderbeine verliert. Dies würde es im Vergleich zu den anderen Mitgliedern seiner Art benachteiligen. Nach den Gesetzen der Evolutionstheorie würde die natürliche Selektion diese behinderte Kreatur bald eliminieren.
Biophysikalische Forschungen haben ergeben, dass Mutationen Veränderungen sind, die sehr selten auftreten. Es ist also unmöglich, dass ein behindertes Tier Millionen Jahre auf die durch Mutationen bedingte komplette Entwicklung seiner Flügel warten könnte, insbesondere, wenn diese Mutationen im Zeitablauf schädigende Effekte haben...
Vögel und Dinosaurier
Die Evolutionstheorie sagt, Vögel hätten sich aus fleischfressenden Theropoden entwickelt. Ein Vergleich zwischen Vögeln und Reptilien zeigt jedoch deren völlig unterschiedliche Eigenschaften, die es unwahrscheinlich machen, dass die einen sich aus den anderen entwickelt haben.
Es gibt diverse strukturelle Unterschiede zwischen Vögeln und Reptilien, einer davon betrifft den Knochenbau. Aufgrund ihrer Größe und des damit verbundenen Gewichts hatten Saurier – die angeblichen Vorfahren der Vögel – dicke, starke Knochen. Vögel dagegen, ob lebend oder ausgestorben, haben hohle Knochen, die sehr leicht sind, damit das entsprechende Tier fliegen kann.
Ein weiterer Unterschied zwischen Reptilien und Vögeln liegt in ihrem Stoffwechselsystem. Reptilien weisen den langsamsten Stoffwechsel des Tierreichs auf. Die Behauptung, Dinosaurier hätten einen warmblütigen, schnellen Stoffwechsel gehabt, bleibt bloße Spekulation. Vögel hingegen befinden sich, was den Stoffwechsel angeht, am anderen Ende der Skala. Reptilien fehlt auf der anderen Seite die Fähigkeit, ihre Körpertemperatur zu regulieren, eben weil sie Kaltblüter sind. Daher setzen sie sich dem Sonnenlicht aus, um sich aufzuwärmen, was wiederum ihren Stoffwechsel in Gang bringt. Einfach gesagt: Reptilien verbrauchen von allen Tieren am wenigsten Energie und Vögel am meisten.
Einer der bekanntesten Ornithologen der Welt, Alan Feduccia von der Universität von North Carolina in den USA, widerspricht der Theorie der Verwandtschaft zwischen Vögeln und Dinosauriern, obwohl er selbst Evolutionist ist. Feduccia sagt folgendes zur Reptil-Vogel Evolution:
Nun, ich habe seit 25 Jahren Vogelschädel studiert, und ich sehe keine wie auch immer geartete Ähnlichkeit. Ich sehe sie einfach nicht... Die Theropoden-Herkunft der Vögel wird meiner Meinung nach zur größten paläontologischen Peinlichkeit des 20. Jahrhunderts werden.108
Larry Martin, ein Spezialist für prähistorische Vögel von der Universität von Kansas, widerspricht ebenfalls der Theorie, dass die Vögel von Sauriern abstammen. Über die Widersprüche, in die sich die Evolutionstheorie bei diesem Thema verstrickt hat, sagt er folgendes:
Um die Wahrheit zu sagen, wenn ich die Dinosaurierherkunft der Vögel mit so wenigen Merkmalen unterstützen sollte, es wäre mir jedes Mal peinlich, wenn ich aufstehen und darüber sprechen sollte.109
DAS EINZIGARTIGE SKELETT DER VÖGEL
Im Gegensatz zu Saurier- und Reptilienknochen sind Vogelknochen hohl. Das verleiht dem Körper Stabilität trotz geringen Gewichts. Die Prinzipien des Vogelskeletts werden beim Entwurf von Flugzeugen, Brücken und anderer Konstruktionen angewandt.

Dinosaurierknochen sind dick und stark aufgrund ihrer massiven Struktur, wogegen die Knochen lebender und ausgestorbener Vögel hohl und deshalb sehr leicht sind.
Gleichwohl, trotz aller Forschungsergebnisse der Wissenschaft wird das haltlose Szenario der Dinosaurier-Vogel Evolution hartnäckig verteidigt. Populäre Publikationen stehen diesem Szenario besonders wohlwollend gegenüber und Konzepte ohne jegliche Beweiskraft werden als Beweise für die Vogel-Dinosaurier Evolution präsentiert. In manchen evolutionistischen Veröffentlichungen werden beispielsweise die unterschiedlichen Hüftknochen verschiedener Saurierarten betont, um die These zu unterstützen, dass Vögel von Sauriern abstammen. Diese so genannten Unterschiede existieren zwischen Saurischia, einer reptilähnlichen Saurierart und Ornithischia, einer vogelähnlichen Art. Der im Beckengürtel befindliche Schambeinknochen der Saurischia zeigt nach vorne, während der der Ornithischia nach hinten zeigt. Diese Ausprägung wird ab und an herangezogen um die Saurier-Vogel Verwandtschaft zu belegen. Doch dafür ist diese anatomische Besonderheit völlig irrelevant. Denn viele andere anatomische Merkmale der Ornithischia weisen überhaupt keine Ähnlichkeit mit Vögeln auf. Zum Beispiel ist der Ankylosaurus ein Dinosaurier, der als Ornithischium klassifiziert wird, mit kurzen Beinen, einem riesigen Körper, bedeckt mit panzerähnlichen Schuppen. Auf der anderen Seite ist der Struthiomimus, der vielmehr den Vögeln ähnelt mit seinen langen Hinterbeinen, kurzen Vorderbeinen und glatter Hautstruktur, tatsächlich ein Saurischium.110
In Kürze, die Anatomie der Hüfte sagt nichts aus und ist kein Beweis für eine evolutionäre Beziehung zwischen Vögeln und Sauriern. Die Behauptung, Saurier ähnelten Vögeln aufgrund der Ähnlichkeit ihrer Hüftknochen ignoriert andere signifikante anatomische Differenzen zwischen den beiden Arten, die jedwede Behauptung einer evolutionären Verbindung, auch vom evolutionistischen Standpunkt gesehen, unhaltbar machen.
Die einzigartige Anatomie der Vogellungen
Ein anderer Faktor, der eindrucksvoll die Unmöglichkeit des Reptil-Vogel Evolutionsszenarios belegt, ist die Anatomie der Lungen von Vögeln, die durch Evolution nicht erklärbar ist.
Bei Landlebewesen zirkuliert die Atemluft bidirektional. Beim Einatmen strömt die Luft durch die Bronchien und gelangt in kleine Luftsäcke, die Alveolen. In ihnen findet der Austausch von Kohlendioxid mit Sauerstoff statt. Danach strömt die verbrauchte Luft beim Ausatmen auf demselben Weg in Gegenrichtung in die Atmosphäre zurück.
Bei Vögeln jedoch fließt die Atemluft nur in eine Richtung. Sie strömt durch eine ausschließlich dem Einatmen dienende Röhre in die Lunge ein und verlässt sie durch eine nur der Ausatmung dienende Röhre. Auf diese Weise können Vögel einen konstanten Einatmungsvorgang aufrechterhalten, ohne diesen zum Ausatmen unterbrechen zu müssen. Dadurch wird die für ihren hohen Energiedurchsatz notwendige Sauerstoffversorgung sichergestellt. Der Molekularbiologe Michael Denton von der Universität von Otago in Neu Seeland erklärt dieses hochspezialisierte Atmungssystem in seinem Buch A Theory in Crisis (Eine Theorie in der Krise):
REPTILIENLUNGE
VOGELLUNGE
1 - Luftstrom, 2 - Alveolen, 3 -Bronchien, 4 -ausströmende Luft, 5 - Parabronchien, 6 - einströmende Luft
Vogellungen funktionieren völlig anders, als die von nicht flugfähigen Tieren. Diese atmen durch dieselben Atemwege ein und aus. Die Vogellunge jedoch wird von einem kontinuierlichen Luftstrom in einer einzigen Richtung durchströmt. Ermöglicht wird dies durch besondere, überall in der Lunge befindliche Luftsäcke. Dank dieses Systems, dessen Details auf der nächsten Seite dargestellt sind, atmen Vögel permanent ein. Dieses Design findet sich ausschließlich bei Vögeln, die während des Fluges einen sehr hohen Sauerstoffdurchsatz haben. Diese Struktur kann sich unmöglich aus Reptilienlungen entwickelt haben, denn keine Kreatur mit einer "Zwischenform" dieser beiden Lungentypen wäre in der Lage, zu atmen.
Bei Vögeln teilt sich die Hauptbronchie in kleine Röhren, die das gesamte Lungengewebe durchdringen. Diese so genannten Parabronchien vereinigen sich später wieder und bilden ein wirkliches Kreislaufsystem, so dass die Luft nur in einer Richtung durch die Lungen fließt... Diese Anatomie der Lunge bei Vögeln ist einzigartig. Es gibt kein anderes Wirbeltier, dessen Lunge dem Vogelsystem nahe kommt. Außerdem ist es bei allen Vögeln in seinen wesentlichen Details identisch, ob beim beim Kolibri, beim Strauss oder beim Falken.111
Der springende Punkt ist, dass die Reptilienlunge mit ihrem bidirektionalen Luftstrom sich nicht zu einer Vogellunge mit unidirektionalem Luftstrom entwickelt haben kann, weil eine Lungenform die "dazwischen" läge, nicht denkbar ist. Damit ein Tier leben kann, muss es konstant atmen und eine Umkehrung der Lungenfunktion würde wegen der dazu notwendigen Änderung der anatomischen Struktur unausweichlich zum Tod führen. Nach der Evolutionstheorie müsste diese Änderung in kleinen Schritten über Millionen Jahre hinweg erfolgen, doch ein Tier dessen Lungen nicht funktionieren, stirbt innerhalb weniger Minuten.
Michael Denton stellt fest, dass es unmöglich ist, eine evolutionistische Erklärung für die Vogellungen zu geben.
Wie ein solches völlig unterschiedliches System sich schrittweise aus der Standardanatomie der Wirbeltiere entwickelt haben soll, ist extrem schwer vorstellbar, besonders wenn man sich klar macht, dass die Aufrechterhaltung der Atmungsfunktion derart lebensnotwendig ist, dass die geringste Fehlfunktion in Minuten zum Tode führt. So wie auch die Federn nicht als Flugorgan funktionieren können, solange nicht ihre Haken und Widerhaken so aneinander angeglichen sind, dass sie perfekt zusammen passen, kann auch die Vogellunge nicht als Atmungsorgan funktionieren, bis dass das sie durchdringende parabronchiale System und das System der Luftsäcke, die den Luftbedarf der Parabronchien sicherstellen, so weit entwickelt sind, dass sie perfekt funktionieren und zusammen arbeiten.112
Kurz, der Übergang von einer terrestrischen zu einer aviatischen Lunge ist unmöglich, weil eine Übergangsform zwecklos wäre.
Ein weiterer Punkt, der hier erwähnt werden muss: Reptilien haben ein Atmungssystem, dass von einem Zwerchfell "bewegt" wird, Vögel haben dagegen ein abdominales Luftsacksystem. Auch dieser anatomische Unterschied macht eine Evolution von einem Lungentypus zum anderen unmöglich, wie John Ruben, eine anerkannte Autorität auf dem Gebiet der Respirationsphysiologie in folgendem Absatz darlegt:
Die frühesten Stadien des Übergangs von einem Vorfahren mit Zwerchfellatmung zum aviatischen abdominalen Luftsacksystem hätten für eine Übergangsform zwischen Theropoden und Vögeln einen Leistenbruch erforderlich gemacht. Solch eine debilisierende Kondition hätte sofort den gesamten Lungenventilationsapparat kompromittiert, und es ist sehr unwahrscheinlich, dass daraus irgendein Selektionsvorteil erwachsen wäre.113
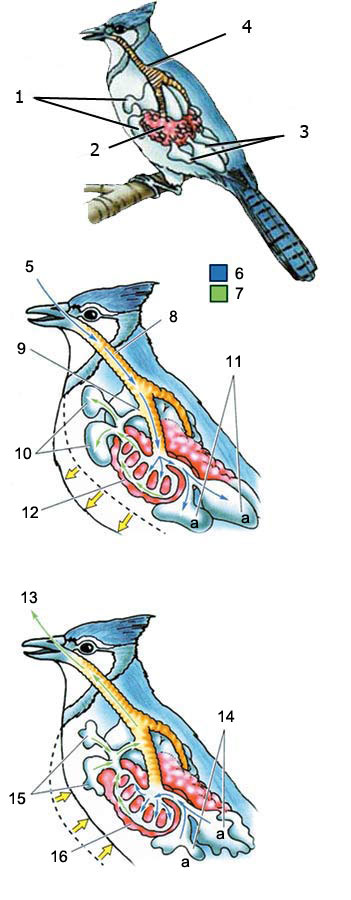
1 - Vordere Luftsäcke, 2 - Lunge, 3 - Hintere Luftsäcke, 4 - Trachea
DAS BESONDERE ATMUNGSSYSTEM DER VÖGEL
EINATMEN: Die Luft, die in das Atmungssystem des Vogels eintritt, gelangt in die Lungen und in hinter ihnen befindliche Luftsäcke. Die verbrauchte Luft strömt in die vorderen Luftsäcke.
AUSATMEN: Wenn ein Vogel ausatmet, gelangt die frische Luft aus den hinteren Luftsäcken in die Lunge. Dieses System stellt einen konstant in die Lunge fließenden Luftstrom sicher.
Die beiden Schaubilder zeigen eine stark vereinfachte Darstellung der vielen Details dieses Lungensystems. Es gibt zum Beispiel spezielle Ventile dort, wo die Luftsäcke mit den Lungen verbunden sind, damit die Luft in die richtige Richtung strömt. All dies zeigt, dass hier Planung am Werk war. Dieses Design erschüttert die Evolutionstheorie nicht nur, sondern es ist der klare Beweis der Schöpfung.
Verbrauchte Luft
Frischluft strömt aus den hinteren Luftsäcken in die Lunge. Verbrauchte Luft wird aus den vorderen Luftsäcken ausgestoßen.
Ein weiteres interessantes Strukturdetail der aviatischen Lunge ist die Tatsache, dass sie niemals luftleer ist und somit nie die Gefahr besteht, dass sie in sich zusammenfällt. Michael Denton erklärt dies:
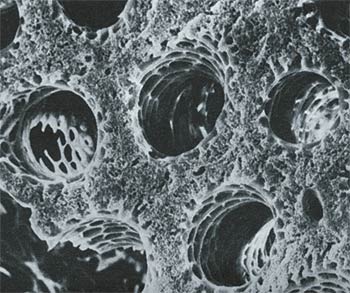
Parabronchiale Röhren, die den Luftstrom in der Vogellunge in die richtige Richtung lenken. Jede dieser Luftröhren hat nur 0,5 Millimeter Durchmesser.
Wie ein solches völlig unterschiedliches Atmungssystem sich schrittweise aus der Standardanatomie der Wirbeltiere entwickelt haben soll, ist wiederum extrem schwer vorstellbar, besonders wenn man sich klar macht, dass die Aufrechterhaltung der Atmungsfunktion für den Organismus absolut lebensnotwendig ist. Außerdem erfordern die einzigartige Funktion und Form der aviatischen Lunge eine Reihe besonderer Anpassungen während ihrer Entwicklung. Wie H.R. Dunker erklärt, eine der Autoritäten der Welt auf diesem Gebiet, ist die aviatische Lunge erstens fest an der inneren Körperwand angewachsen und kann sich infolgedessen nicht zwecks Volumenvergrößerung ausdehnen, zweitens kann sie wegen des geringen Durchmessers der Lungenkapillaren und der daraus resultierenden hohen Oberflächenspannung jeder in ihnen befindlichen Flüssigkeit aus einem zusammengesunkenen Zustand nicht aufgefüllt werden, wie dies bei allen anderen Wirbeltieren von Geburt an geschieht. Die Luftkapillaren ziehen sich niemals zusammen, wie die Alveolen anderer Wirbeltiere. Die Parabronchien sind in das Lungengewebe hineingewachsene, offene Röhren, die entweder mit Wasser oder mit Luft gefüllt sind.114
Mit anderen Worten, die Atemwege in den Lungen der Vögel sind so eng, dass die Luftsäcke in ihren Lungen sich nicht mit Luft füllen und sich wieder entleeren können, wie dies bei Landtieren der Fall ist.
Würde eine Vogellunge sich jemals komplett entleeren, könnte der Vogel sie nicht wieder mit Luft füllen. Aus diesem Grund ermöglichen die überall in der Lunge befindlichen Luftsäcke einen konstanten Luftstrom, der die Lungen vor Entleerung schützt.
Natürlich kann dieses System, das sich vollständig von Reptilienlungen und den Lungen anderer Wirbeltiere unterscheidet und das auf einem höchst sensiblen Gleichgewicht basiert, niemals durch ohne Bewusstsein erfolgende Mutationen zustande gekommen sein, schrittweise, wie uns die Evolutionstheorie weismachen will. Denton beschreibt die aviatische Lunge folgendermaßen, was den Darwinismus erneut widerlegt:
Die aviatische Lunge bringt uns nahe an die Antwort auf Darwins Herausforderung: "Wenn gezeigt werden könnte, dass irgendein komplexes Organ existiert, das nicht durch zahlreiche, aufeinanderfolgende, leichte Modifikationen entstanden ist, würde meine Theorie vollständig zusammenbrechen." 115Vogelfedern und Reptilienschuppen
Eine andere unüberwindliche Kluft zwischen Vögeln und Reptilien wird durch die Federn gebildet, deren Besitz einzig den Vögeln vorbehalten ist. Die Körper der Reptilien sind mit Schuppen bedeckt, die der Vögel mit Federn. Die Hypothese, dass Vogelfedern sich aus Reptilienschuppen entwickelt hätten, ist völlig unbegründet und wird durch den Fossilienbestand widerlegt, wie die Paläontologin Barbara Stahl zugeben muss:
Wie (Federn) nun ursprünglich entstanden sind, vermutlich aus Reptilienschuppen, entzieht sich der Analyse... Es scheint, dass die Evolution von Reptilienschuppen zu der komplexen Konstruktion von Federn einen immensen Zeitraum in Anspruch genommen haben muss und sie muss über eine Serie von Übergangsformen erfolgt sein. Bis jetzt gibt der Fossilienbestand keinen Grund zu dieser Annahme.116
A. H. Brush, Professor für Physiologie und Neurobiologie an der Universität von Connecticut, sieht diese Realität ein, obwohl er selbst Evolutionist ist:
"Jedes einzelne Merkmal, von der Genstruktur und Genorganisation zur Entwicklung, Morphogenese bis zum Gewebeaufbau ist unterschiedlich (bei Federn und Schuppen).117
Professor Brush untersucht auch die Proteinstruktur von Vogelfedern und stellt fest, sie sind "einzigartig unter den Wirbeltieren".118
Es gibt nichts an Fossilien, das beweisen würde, dass sich Vogelfedern aus Reptilienschuppen entwickelt hätten. Im Gegenteil, Federn tauchen plötzlich im Fossilienbestand auf, wie Professor Brush bemerkt, als ein einzigartiges Charakteristikum der Vögel.119
Nebenbei bemerkt, bisher wurde kein epidermes Gewebe entdeckt, dass ein Ausgangspunkt für Vogelfedern hätte sein können.120
Schon viele Fossilien sind bereits Gegenstand von Spekulationen über gefiederte Dinosaurier gewesen, doch genaue Untersuchungen erwiesen jedes Mal das Gegenteil. Der prominente Ornithologe Alan Feduccia schreibt folgendes in seinem Artikel "Warum den Sauriern die Federn fehlten":
Federn sind einzigartig für Vögel und es gibt keine bekannten Übergangsformen zwischen Reptilienschuppen und Federn. Ungeachtet der Spekulationen über die Natur der gestreckten Schuppen bei Formen wie Longisquama... dass diese federähnliche Strukturen seien, gibt es einfach keinen vorzeigbaren Beweis, dass sie es tatsächlich sind.121

Das Sinosauropteryx Fossil, von evolutionistischen Paläontologen als gefiederter Dinosaurier vorgestellt, was sich als völliger Unfug erwies.

REPTILIENSCHUPPEN
Die Schuppen, die den Reptilienkörper bedecken, unterscheiden sich vollständig von Vogelfedern. Im Gegensatz zu Federn erstrecken sich die Schuppen nicht unter die Haut, sondern bilden eine harte Schicht auf der Oberfläche des Tierkörpers. Weder genetisch, noch biochemisch oder anatomisch haben Schuppen Ähnlichkeiten mit Federn. Der große Unterschied zwischen ihnen zeigt wieder einmal, dass das Szenarium der Evolution von Reptilien zu Vögeln unbegründet ist.
Die Anatomie der Federn
Der anatomische Aufbau und das gesamte Design der Vogelfedern sind so komplex, dass dieses Phänomen niemals als Resultat eines Evolutionsprozesses erklärt werden kann. Die Feder besteht aus einem in der Haut verwurzelten Schaft, an dem sich an zwei gegenüberliegenden Seiten wimpernähnliche, mit hunderten winziger Haken versehene Haare befinden. Diese Haken dienen dem Zusammenhalt der Haare, wodurch eine flexible und doch stabile Struktur entsteht. Bricht diese Struktur aus irgendeinem Grund auf, entsteht ein Spalt zwischen den Haaren, den der Vogel jedoch mühelos "reparieren" kann, indem er sich schüttelt oder die betroffene Stelle mit seinem Schnabel glattstreicht. Das nach dem Reißverschlussprinzip funktionierende Hakensystem an den Federhaaren sorgt dafür, dass jedes einzelne Haar in seine Ausgangsposition zurückkehrt.
Zu behaupten, das komplexe Design der Federn könnte durch die Evolution von Reptilienschuppen per Zufallsmutationen entstanden sein, ist nichts als ein dogmatischer Glaube ohne jegliche wissenschaftliche Grundlage. Selbst einer der Wortführer des Darwinismus, Ernst Mayr, machte vor einigen Jahren folgendes Eingeständnis:
DIE KOMPLEXE STRUKTUR VON VOGELFEDERN

Wenn man Vogelfedern genau betrachtet, kommt ein sehr filigranes Design zum Vorschein. Da sind noch kleinere Härchen an jedem kleinen Haar, und diese haben Widerhaken, die die Haare miteinander verbinden. Die Bilder zeigen stark vergrößerte Vogelfedern.
Es ist eine erhebliche Strapazierung der Gutgläubigkeit eines Menschen, anzunehmen, dass so fein ausbalancierte Systeme wie die bestimmter Sinnesorgane (das Auge von Wirbeltieren oder die Vogelfeder) durch zufällige Mutationen verbessert werden könnten.122
Das Design der Federn brachte auch Darwin ins Nachdenken. Die perfekte Ästhetik der Pfauenfedern machte ihn nach eigenen Worten "krank". In einem Brief an Asa Gray vom 3. April 1860 schrieb er:
"Ich kann mich gut an die Zeit erinnern wenn der Gedanke an das Auge mich frösteln ließ, aber über dieses Stadium der Beschwerden bin ich hinweg..." und fuhr dann fort: "... und jetzt lassen manche belanglose strukturelle Einzelheiten mich mitunter sehr unwohl fühlen. Der Anblick einer Pfauenschwanzfeder macht mich jedes Mal krank!"123
Kurz, die enormen Strukturunterschiede zwischen Vogelfedern und Reptilienschuppen und das unglaublich komplexe Design von Federn zeigen ganz klar die Haltlosigkeit der Behauptung, dass Federn sich aus Schuppen entwickelt hätten.
Die falschen Vorstellungen über Archäopteryx

Einer der Beweise, dass Archäopteryx ein fliegender Vogel war, ist seine asymmetrische Federnstruktur. Oben: ein Federnfossil von Archäopteryx.
Als Antwort auf die Frage, ob es irgendeinen Beweis gibt für die "Reptil-Vogel Evolution", nennen Evolutionisten den Namen eines einzigen Tieres: Archäopteryx, eine der bekanntesten so genannten Übergangsformen von den wenigen, die die Evolutionisten immer noch als solche behaupten.
Archäopteryx, der von den Evolutionisten so genannte Vorfahr moderner Vögel, lebte vor schätzungsweise 150 Millionen Jahren. Die Theorie besagt, kleine Saurier wie der Velociraptor oder der Dromäosaurier hätten während ihrer Evolution Flügel erworben und zu fliegen begonnen. Daher wird der Archäopteryx als Übergangsform gesehen, die sich von ihren Sauriervorfahren abspaltete und die erste war, die die Fähigkeit zu fliegen entwickelte.
Jüngste Studien an Archäopteryx Fossilien weisen jedoch darauf hin, dass dieser Erklärung jede wissenschaftliche Grundlage fehlt. Es handelt sich auf keinen Fall um eine Übergangsform, sondern um eine ausgestorbene Vogelart, die einige unwesentliche Abweichungen von heutigen Vögeln aufweist.
Die These, Archäopteryx sei nur ein "halber" Vogel gewesen, der nur unbeholfen fliegen konnte, war bis vor kurzem in evolutionistischen Kreisen sehr populär. Das bei diesem Tier angeblich fehlende Sternum (Brustbein) wurde als Hauptargument dafür gebraucht, dass der Vogel nicht perfekt fliegen konnte. Das Sternum ist der Knochen des Brustkorbs, an dem die zum Fliegen notwendigen Muskeln angewachsen sind. Es ist bei allen fliegenden und nicht fliegenden Vögeln vorhanden, sogar bei Fledermäusen, einem fliegenden Säugetier, das einer ganz anderen Familie angehört. Das 1992 gefundene siebente Archäopteryx Fossil jedoch entkräftete dieses Argument, denn es wies eindeutig ein Brustbein auf, das nach Meinung der Evolutionisten nicht existierte. Das Fossil wurde von Nature so beschrieben:
Das kürzlich gefundene siebente Archäopteryx Exemplar weist ein teilweise erhaltenes winkliges Brustbein auf, dessen Existenz schon lange vermutet worden war, jedoch zuvor nie nachgewiesen werden konnte. Dies bestätigt nun auch das Vorhandensein starker Flugmuskeln, doch die Fähigkeit, lange in der Luft bleiben zu können, scheint fraglich.124
Die Entdeckung entwertet natürlich die Behauptung, Archäopteryx sei ein "halber" Vogel gewesen, der nicht richtig fliegen konnte.
Darüber hinaus wurde die Anatomie der Vogelfeder zu einem der wichtigsten Beweise, dass Archäopteryx ein fliegender Vogel im wahrsten Sinn des Wortes war. Seine asymmetrische Federstruktur unterscheidet sich nicht von den modernen Vögeln und weist darauf hin, dass Archäopteryx perfekt fliegen konnte. Wie der berühmte Paläontologe Carl O. Dunbar feststellt: "Wegen seiner Federn (ist Archäopteryx) eindeutig als Vogel zu klassifizieren."125 Paläontologe Robert Carrol erklärt dazu weiter:
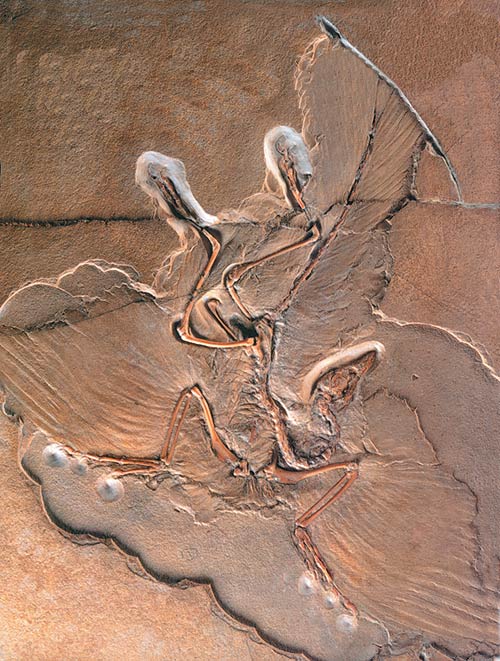
Welche sind auf dem Display in berühmteste Berliner Archaeopteryx fossilen
Die Geometrie der Flugfedern des Archäopteryx ist identisch mit der von modernen fliegenden Vögeln, nicht flugfähige Vögel hingegen haben symmetrische Federn. Die Art und Weise, in der die Federn auf dem Flügel angeordnet sind, entspricht ebenfalls den modernen Vögeln... Nach van Tyne und Berger sind die relative Größe und die Flügelform von Archäopteryx ähnlich wie bei Vögeln, die sich durch enge Öffnungen bewegen müssen, wie hahnenartige Vögel, Tauben, Waldschnepfen, Spechte und die meisten Sperlingsarten... Deren Flugfedern sind seit 150 Millionen Jahren demselben Zustand.126
Eine andere Tatsache, die durch die Archäopteryxfedern aufgedeckt wurde, ist sein warmblütiger Stoffwechsel. Wie schon erwähnt, sind Reptilien und Saurier Kaltblüter, deren Körpertemperatur sich an die Umgebung anpasst, die also nicht homöostatisch reguliert wird. Eine sehr wichtige Funktion der Vogelfedern ist die Aufrechterhaltung einer konstanten Körpertemperatur. Die Tatsache, dass Archäopteryx Federn hatte zeigt, dass er im Gegensatz zu Sauriern ein warmblütiger Vogel war, der seine Körpertemperatur konstant halten musste.
Die Zähne und Klauen von Archäopteryx

Wie der Archäopteryx hat der Vogel Opisthocomus hoazin, der heute lebt, klauenähnliche Nägel an den Flügeln.
Zwei wichtige Punkte, auf denen evolutionistische Biologen herumreiten, wenn sie behaupten, Archäopteryx sei eine Übergangsform gewesen, sind die Klauen an seinen Flügeln und ihre Zähne.
Es ist wahr, dass Archäopteryx Zähne und Klauen hatte, doch diese Eigenschaften implizieren nicht automatisch eine Verwandtschaft mit Reptilien. Im übrigen gibt es zwei heute lebende Vogelarten, Touraco und Hoatzin, die über Klauen verfügen, mit denen sie sich an Zweigen festhalten. Diese Kreaturen sind Vögel und nichts anderes, sie haben keinerlei Reptilieneigenschaften. Daher ist es völlig haltlos, darauf zu bestehen, Archäopteryx nur wegen seiner Klauen an den Flügeln als eine Übergangsform zu bezeichnen.
Auch die Zähne im Schnabel des Archäopteryx implizieren keineswegs, dass er eine Übergangsform ist. Evolutionisten liegen falsch, wenn sie behaupten, diese Zähne seien charakteristisch für Reptilien, denn Zähne sind kein typisches Merkmal für Reptilien. Manche heute lebenden Reptilien haben Zähne, andere nicht. Archäopteryx ist Außerdem nicht die einzige Vogelart, die Zähne besaß. Es stimmt zwar, dass es heute keine Vögel mit Zähnen mehr gibt, doch wenn wir den Fossilienbestand anschauen, sehen wir, dass zu Lebzeiten von Archäopteryx und danach - sogar bis vor kurzem – eine ganze Gruppe verschiedener Vogelarten existiert hat, die Zähne aufwies.
Der wichtigste Punkt aber ist, dass die Zahnanatomie von Archäopteryx und anderen "Zahnvögeln" völlig abweicht von der ihrer so genannten Vorfahren, den Sauriern. Die bekannten Ornithologen L. D. Martin, J. D. Stewart, und K. N. Whetstone beobachteten, dass Archäopteryx und ähnliche Vögel ungezackte Zähne mit schmalen Zahnhälsen und ausgedehnten Wurzeln haben. Die Zähne der theropodischen Saurier hingegen, der angeblichen Vorfahren dieser Vögel, hatten gezackte Zähne mit geraden Wurzeln.127 Die Forscher verglichen auch die Fußknochen des Archäopteryx mit denen der Saurier und fanden keinerlei Ähnlichkeit zwischen ihnen.128
Studien von Anatomen wie S. Tarsitano, M.K. Hecht, und A.D. Walker haben ergeben, dass manche Ähnlichkeiten, die John Ostrom und andere bei den Gliedmassen von Archäopteryx und Sauriern gesehen haben, in Wahrheit Fehlinterpretationen waren.129 Zum Beispiel analysierte A.D: Walker die Ohrregion von Archäopteryx und fand heraus, dass sie derjenigen moderner Vögel sehr ähnlich ist.130
J. Richard Hinchliffe, vom Institut für biologische Wissenschaften an der Universität von Wales studierte die Anatomie von Vögeln und deren angeblichen Vorfahren mit Hilfe moderner Isotopen-Techniken und entdeckte, dass Saurier an den Vorderpfoten die drei Finger I, II und III aufweisen, während es bei Vogelflügeln die II, III und IV sind. Dies stellt ein großes Problem dar für die Befürworter der Archäopteryx-Saurier Verbindung.131 Hinchliffe veröffentlichte seine Studien 1997 in Science, wo er schrieb:
Zweifel an der Homologie zwischen theropodischen und Vogelfingern erinnern uns an einige der anderen Probleme der "Saurierursprungs"-Hypothese. Es sind die folgenden: (i) Die viel kleineren (relativ zur Körpergröße) theropodischen vorderen Gliedmassen im Vergleich zum Archäopteryxflügel. Solch kleine Gliedmassen können nicht überzeugen als Herkunft der Flugfähigkeit der relativ schweren Saurier. (ii) Die bei Theropoden seltenen halbmondförmigen Handgelenkknochen, die nur bei vier Arten bekannt sind (einschließlich dem Deinonychus). Die meisten Theropoden haben relativ viele Handgelenk-Elemente, die schwer mit denen von Archäopteryx zu homologisieren sind. (iii) Das Zeitparadoxon, dass die meisten theropodischen Saurier und besonders die vogelähnlichen Dromäosaurier erst sehr viel später im Fossilienbestand auftauchen als Archäopteryx.
Wie Hinchliffe bemerkt, ist das Zeitparadoxon einer der Fakten, der die evolutionistischen Behauptungen über Archäopteryx obsolet macht. Der bereits im ersten Kapitel erwähnte amerikanische Molekularbiologe Jonathan Wells schreibt in Ikonen der Evolution dass Archäopteryx zu einer Ikone der Evolutionstheorie gemacht worden ist, obwohl es klare Beweise dafür gibt, dass er nicht der primitive Vorfahr der Vögel ist. Wie soll dies auch möglich sein, wenn theropodische Saurier – die angeblichen Vorfahren – jünger sind als Archäopteryx? "Zweibeinige Reptilien, die auf dem Boden umherliefen und noch andere Eigenschaften hatten, die man von einem Vorfahren des Archäopteryx erwarten dürfte, erscheinen erst später.132
All diese Erkenntnisse weisen darauf hin, dass Archäopteryx keine Übergangsform war, sondern nur ein Vogel, den man in die Kategorie "Zahnvögel" einordnen muss. Dieses Tier mit theropodischen Sauriern in Verbindung bringen zu wollen, ist völlig unsinnig. In einem Artikel überschrieben mit "Der Untergang der Vögel sind Dinosaurier" Theorie schreibt der amerikanische Biologe Richard L. Deem über Archäopteryx und die Behauptung der Vogel-Saurier Evolution:
Die Ergebnisse jüngerer Studien zeigen, dass die Pfoten der theropodischen Saurier sich aus den Fingern I II und III herleiten, während sich die Vogelflügel, obwohl sie strukturell ähnlich aussehen, sich aus den Fingern II, III und IV herleiten... Es gibt noch weitere Probleme mit der "Vögel sind Saurier" Theorie. Die theropodischen Vordergliedmassen sind wesentlich kleiner (relativ zur Körpergröße) als die des Archäopteryx. Der kleine Flügelvorläufer des Theropoden ist nicht sehr überzeugend, besonders wenn man das hohe Gewicht dieser Saurier in Betracht zieht. Der großen Mehrheit der Theropoden fehlt der halbmondförmige Handgelenkknochen und sie haben eine hohe Zahl anderer Handgelenkelemente, die keine Homologie mit den Archäopteryxknochen aufweisen. Außerdem tritt bei fast allen Theropoden der Nerv V1 seitlich zusammen mit anderen Nerven aus dem Schädelknochen aus, während er bei Vögeln durch eine eigene Öffnung aus der Vorderseite des Schädels austritt. Weiterhin gibt es das kleine Problem, dass die große Mehrheit der Theropoden erst nach dem Auftauchen von Archäopteryx erscheint.133
Archäopteryx und andere Urvogel-Fossilien
Einige kürzliche entdeckte Fossilien entwerten das evolutionistische Szenario auch in anderem Zusammenhang.

Der Confuciusornis, der zur gleichen Zeit wie Archäopteryx lebte, weist viele Ähnlichkeiten mit modernen Vögeln auf.
Lianhai Hou und Zhonghe Zhou, zwei Paläontologen am chinesischen Institut für Wirbeltierpaläontologie entdeckten 1995 ein neues Vogelfossil und nannten es Confuciusornis. Es hat fast dasselbe Alter wie Archäopteryx, etwa 140 Millionen Jahre, aber keine Zähne im Mund. Schnabel und Federn zeigen dieselben Eigenschaften, wie heutige Vögel. Confuciusornis hat dieselbe Skelettanatomie wie moderne Vögel, aber auch Klauen an den Flügeln, genau wie Archäopteryx. Eine andere Struktur, die den Vögeln eigen ist und die Schwanzfedern unterstützt, fand sich auch bei Confuciusornis.134 In Kürze, dieses Fossil, dass genauso alt ist wie Archäopteryx, den man zunächst für den ersten Vogel hielt und als halbes Reptil ansah, sieht fast genauso aus wie ein moderner Vogel. Diese Tatsache hat alle evolutionistischen Behauptungen widerlegt, Archäopteryx sei der primitive Vorfahr aller Vögel.
Ein anderes in China ausgegrabenes Fossil sorgte für noch größere Verwirrung. Im November 1996 gaben L. Hou, L. D. Martin, und Alan Feduccia in Science die Existenz eines 130 Millionen Jahre alten Vogels namens Liaoningornis bekannt, Er hatte ein Brustbein, an dem die Flugmuskeln angewachsen waren, genau wie bei modernen Vögeln.135 Der Vogel war auch in anderen Einzelheiten nicht von modernen Vögeln zu unterscheiden. Der einzige Unterschied waren seine Zähne. Dies zeigte, dass Vögel mit Zähnen nicht die von Evolutionisten angenommenen primitiven Strukturen besaßen. Ein Artikel in Discover stellte fest, dass Liaoningornis die Merkmale moderner Vögel aufwies: "Woher kamen die Vögel? Das Fossil legt nahe, dass es nicht von Dinosauriern abstammt."136
Ein anderes Fossil, dass die evolutionistischen Behauptungen über Archäopteryx widerlegt, ist der Eoalulavis. Seine Flügelanatomie, die 25 bis 30 Millionen Jahre jünger als die des Archäopteryx sein soll, ist auch bei modernen, langsam fliegenden Vögeln zu beobachten.137 Das beweist, dass vor 120 Millionen Jahren Vögel am Himmel flogen, die von modernen Vögeln in vielerlei Hinsicht nicht zu unterscheiden waren.
Diese Fakten besagen einmal mehr, dass weder Archäopteryx noch andere ähnliche Urvögel Übergangsformen gewesen sind. Die Fossilien geben keinen Hinweis darauf, dass verschiedene Vogelarten sich voneinander entwickelt haben. Im Gegenteil, der Fossilienbestand beweist, dass heutige moderne Vögel und einige archaische Arten wie der Archäopteryx tatsächlich gleichzeitig auf der Erde lebten. Es ist wahr, dass einige dieser Arten inzwischen ausgestorben sind, doch nur die Tatsache, dass einige der Arten, die einmal existiert haben, bis auf den heutigen Tag überleben konnten, beweist nicht die Evolutionstheorie.
Archäoraptor: Der Dino-Vogel Schwindel
National Geographic dergisinin kuşların evrimi senaryosunun delili olarak tanıttığı Archaeoraptor adlı "dino-kuş"un, bir yıl sonra sahte bir fosil olduğu ortaya çıktı.
Nachdem ihnen nicht gelungen war, in Archäopteryx das zu finden, wonach sie gesucht hatten, setzten die Advokaten der Evolutionstheorie in den 1990er Jahren ihre Hoffnungen auf andere Fossilien, und eine ganze Serie so genannter Dino-Vogel Fossilien tauchte in den Medien der Welt auf. Doch schnell wurde entdeckt, dass diese einfach Fehlinterpretationen waren oder schlimmer noch, Fälschungen.
Das erste Dino-Vogel Märchen war die Geschichte des in China ausgegrabenen "gefiederten Dinosaurier-Fossils", die 1996 mit großem Medienaufwand vorgestellt wurde. Ein Reptilienfossil namens Sinosauropteryx war gefunden worden, und einige Paläontologen, die das Fossil untersuchten, sagten, es habe im Gegensatz zu modernen Reptilien Vogelfedern gehabt. Ein Jahr später durchgeführte Untersuchungen zeigten jedoch, dass das Fossil in Wahrheit keinerlei Struktur aufwies, die einer Vogelfeder ähnelt. Ein Artikel in Science mit dem Titel "Der gefiederte Dinosaurier wird gerupft" führte aus, dass die von evolutionistischen Paläontologen für Federn gehaltenen anatomischen Besonderheiten definitiv nichts zu tun haben mit Vogelfedern:
Vor genau einem Jahr waren Paläontologen in heller Aufregung über Photos eines so genannten gefiederten Dinosauriers, die auf dem Jahrestreffen der Gesellschaft für Wirbeltierpaläontologie herumgereicht wurden. Der Sinosauropteryx aus der Yixian Formation in China brachte es auf die Titelseite der New York Times und wurde von vielen als Bestätigung dafür angesehen, dass Vögel von Sauriern abstammen. Doch auf dem diesjährigen Jahrestreffen der Wirbeltierpaläontologen in Chikago im vergangenen Monat fiel das Urteil etwas anders aus: Die fraglichen Strukturen sind keine Federn, ist sich das halbe Dutzend Paläontologen einig, die die Photos zu sehen bekommen haben. ...Larry Martin von der Universität des Staates Kansas in Lawrence hält die Strukturen für aufgescheuertes Gewebe unter der Haut, das infolgedessen nichts zu tun hat mit Vogelfedern.138
Eine noch größere Dino-Vogel Sensationshysterie brach 1999 aus. In seiner Novemberausgabe veröffentlichte National Geographic einen Artikel über ein in China ausgegrabenes Fossil, dass angeblich Eigenschaften von Vögeln und Sauriern aufwies. Christopher P. Sloan, der Autor des Artikels, ging so weit zu behaupten: "Wir können nun mit derselben Sicherheit sagen, dass Vögel Theropoden sind, wie wir sagen dass Menschen Säugetiere sind." Diese Spezies, die vor 125 Millionen Jahren gelebt haben soll, erhielt sofort den Namen Archaeoraptor liaoningensis.139
Doch das Fossil war eine Fälschung, die kunstvoll aus den Überresten von fünf verschiedenen Fossilien zusammengesetzt worden war. Eine Gruppe von Wissenschaftlern, unter denen drei Paläontologen waren, bewies die Fälschung ein Jahr später mit Hilfe eines Computertomographen und war in Wahrheit das Werk eines chinesischen Evolutionisten. Der Dino-Vogel war mit Hilfe von Leim, Zement aus 88 Knochen und Steinen zusammengesetzt. Forschungen weisen darauf hin, dass Archaeoraptor liaoningensis aus dem Vorderteil des Skeletts eines Urvogels sowie aus Teilen von vier anderen Fossilien zusammengebastelt wurde.
Das Interessante daran ist, dass National Geographic einen Schwerpunktartikel über eine so grobe Fälschung brachte und diese Fälschung als Basis für die Behauptung benutzte, dass die Szenarien der Vogelevolution sich nun bestätigt hätten, ohne dass in dem Artikel der leiseste Zweifel daran geäußert wurde. Dr. Storrs Olson, Leiter des berühmten Nationalen Museums für Naturgeschichte des Smithsonian Instituts in den USA sagte später, er habe das National Geographic zuvor gewarnt, dass Fossil sei eine Fälschung, doch das Management des Magazins habe ihn völlig ignoriert. Olson zufolge "hat das National Geographic inzwischen das allerniedrigste Niveau erreicht, indem es einen substanzlosen, sensationslüsternen Boulevardjournalismus betreibt."140
In einem Brief an Peter Raven von National Geographic beschreibt Olson sehr detailliert die wahre Geschichte der Hysterie über den "gefiederten Dinosaurier" seitdem sie in einem vorangegangenen Artikel des National Geographic aus dem Jahr 1998 zum ersten Mal erschienen war:
Vor der Veröffentlichung des Beitrags "Dinosauriern wachsen Flügel" in der Juli-Ausgabe 1998 des National Geographic lud mich Lou Mazzatenta, der Photograph für Sloan's Artikel in die Redaktion ein, um mich die Photos der chinesischen Fossilien anschauen zu lassen und meine Meinung über den Tenor des beabsichtigen Artikels zu hören. Ich wies damals darauf hin, dass es von dem zur Veröffentlichung vorgesehenen Standpunkt des National Geographic erheblich abweichende Meinungen gebe, die breite Unterstützung fänden, doch später wurde mir klar, dass das National Geographic an nichts anderem interessiert war, als an der vorherrschenden dogmatischen Ansicht, dass sich Vögel aus Sauriern entwickelt hätten.
Sloan's Artikel hebt die Voreingenommenheit auf ein ganz neues Niveau, er besteht zum großen Teil aus nicht verifizierbaren oder undokumentierten Informationen, die die Neuigkeit eigentlich erst erzeugen, anstatt über sie zu berichten. Seine gewagte Behauptung, "Wir können nun mit derselben Sicherheit sagen, dass Vögel Theropoden sind, wie wir sagen, dass Menschen Säugetiere sind", versucht noch nicht einmal den Eindruck zu erwecken, als handele es sich um die Ansicht eines bestimmten Wissenschaftlers oder einer Gruppe von Wissenschaftlern, sie ist praktisch nichts als die Privatpropaganda eines Redakteurs. Diese melodramatische Behauptung war bereits zuvor durch embryologische und vergleichende morphologische Studien widerlegt worden, die selbstverständlich nicht erwähnt werden.
Wichtiger ist jedoch die Tatsache, dass keine der in Sloan's Artikel abgebildeten Strukturen, die Federn darstellen sollen, auch nachgewiesenermaßen Federn sind. Zu behaupten, sie seien es, ist wenig mehr als Wunschdenken, das als Tatsache präsentiert wird. Die Erklärung auf Seite 103, dass "hohle, haarähnliche Strukturen diese Protofedern charakterisieren" würden, ist Unsinn, da Protofedern nur als theoretisches Konstrukt existieren, so dass deren interne Struktur völlig hypothetisch ist.
Die Sensationsmache über gefiederte Dinosaurier in der Ausstellung, die kürzlich in der National Geographic Society gezeigt wurde, ist noch haltloserer Natur, sie stellt die völlig aus den Fingern gesogene Behauptung auf, es gebe stichhaltige Beweise, dass eine Vielzahl fleischfressender Saurier ein Federkleid gehabt hätte. Ein Modell des Sauriers Deinonychus und Abbildungen eines Baby-Tyrannosaurus Rex werden mit Federn bedeckt gezeigt, das alles ist reine Phantasie und hat außerhalb von Science Fiction nichts zu suchen.
Mit freundlichen Grüssen
Storrs L. Olson
Nationales Museum für Naturgeschichte
Smithsonian Institut 141
Dieser Fall zeigt zwei wichtige Tatsachen. Erstens, es gibt Menschen, die keinerlei Bedenken haben, zum Mittel der Fälschung zu greifen, um die Evolutionstheorie zu beweisen. Zweitens, einige höchst respektierte populärwissenschaftliche Magazine, deren Mission es ist, den Menschen die Evolutionstheorie aufzuzwingen, sehen kein Problem darin, bewusst alle Fakten zu missachten, die ihnen unbequem sind oder alternative Interpretationen zulassen. So sind diese Magazine zu Propagandawerkzeugen der Evolutionstheorie geworden. Sie nehmen nicht wissenschaftlich Stellung, sondern dogmatisch, und sie kompromittieren ganz bewusst die Wissenschaft, um die Evolutionstheorie zu verteidigen, der sie dermaßen verfallen sind.
Ein weiterer wichtiger Aspekt ist die Tatsache, dass es keinerlei Beweis gibt für die These, Vögel hätten sich aus Dinosauriern entwickelt. Wegen dieses Mangels an Beweisen werden entweder gefälschte "Beweise" produziert oder Fakten werden uminterpretiert. Es gibt in Wahrheit noch nicht einmal einen Beweis dafür, dass sich Vögel überhaupt aus anderen Lebewesen entwickelt haben. Dagegen zeigen alle bisherigen Entdeckungen, dass Vögel im Vollbesitz aller ihrer charakteristischen Körpermerkmale auf der Erde erschienen sind.
Die Herkunft der Insekten
Im Zusammenhang mit der Herkunft der Vögel haben wir die Lauftheorie erwähnt, die von evolutionistischen Biologen vorgeschlagen wird. Wie wir an dieser Stelle klar gemacht haben, beinhaltet die Antwort auf die Frage, wie den Reptilien Flügel gewachsen sind, die Spekulation darüber, sie hätten "versucht, mit ihren Vorderfüßen Insekten zu fangen". Nach dieser Theorie entwickelten sich die Vorderbeine der Reptilien auf der Jagd nach Insekten mit der Zeit langsam zu Flügeln.

Eine Fliege, vor 35 Millionen Jahren in Bernstein eingeschlossen. Auch dieses an der baltischen Küste gefundene Fossil unterscheidet sich nicht von den Fliegen unserer Zeit


Dieser Acantherpestes major Tausendfüßler, gefunden in Kansas/USA ist mehr als 300 Millionen Jahre alt und unterscheidet sich nicht von heutigen Tausendfüßlern.
Ein 145 Millionen Jahre altes Fliegen-Fossil, gefunden in Liaoning in China. Es gehört zu derselben Art wie die heutigen Fliegen.
Wir haben bereits ausgeführt, dass diese Theorie auf keinerlei wie auch immer gearteten wissenschaftlichen Entdeckungen beruht. Doch es gibt noch eine andere interessante Seite an ihr, die wir noch nicht angesprochen haben. Fliegen können bereits fliegen. Wie sind sie also zu ihren Flügeln gekommen? Allgemein gefragt, woher kommen die Insekten, von denen Fliegen nur eine einzige Klasse sind?
In der Klassifizierung der Lebewesen bilden die Insekten eine Unterabteilung der der Abteilung Arthropoden (Gliederfüßler). Das bisher älteste Insektenfossil stammt aus dem Devon-Zeitalter von vor 410 bis 360 Millionen Jahren. Im darauf folgenden pennsylvanischen Zeitalter von vor 325 bis 286 Millionen Jahren tauchte eine große Zahl verschiedener Insektenarten auf. Zum Beispiel sind die Kakerlaken ganz plötzlich da und zwar mit derselben Anatomie, die sie noch heute haben. Betty Faber vom Amerikanischen Museum für Naturgeschichte berichtet, dass die 350 Millionen Jahre alten versteinerten Kakerlaken die gleichen sind wie die, die heute leben.142 Kreaturen wie Spinnen, Zecken und Tausendfüssler sind keine Insekten, sondern gehören zu anderen Unterabteilungen der Arthropoden. Wichtige Fossilienfunde dieser Kreaturen wurden 1983 auf dem Jahrestreffen der Amerikanischen Vereinigung für den Fortschritt der Wissenschaft bekannt gegeben. Das Interessante an den über 380 Millionen Jahre alten Spinnen-, Zecken- und Tausendfüsslerfossilien ist die Tatsache, dass sie sich überhaupt nicht von den heute lebenden Exemplaren unterscheiden. Einer der Wissenschaftler, der die Fossilien untersuchte, bemerkte, dass "sie aussehen, als seien sie gerade gestern gestorben".143
Auch geflügelte Insekten tauchen plötzlich im Fossilienbestand auf, mit all ihren besonderen Eigenschaften. Zum Beispiel wurde eine große Zahl von Libellenfossilien aus dem pennsylvanischen Zeitalter gefunden. Auch diese Libellen weisen genau dieselbe Anatomie auf, wie ihre Nachfahren heute.

Es gibt keinen Unterschied zwischen diesem 320 Millionen Jahre alten Schaben-Fossil und heute lebenden Schaben.
Libellen und Fliegen tauchen sehr plötzlich auf, zusammen mit flügellosen Insekten. Das widerlegt die Theorie, flügellose Insekten hätten Flügel ausgebildet und sich Schritt für Schritt zu fliegenden Insekten entwickelt. In einem Kapitel ihres Buches Biomechanik in der Evolution, schreiben Robin Wootton und Charles P. Ellington dazu folgendes:
Als in der mittleren und jüngeren Karbonzeit die ersten Insektenfossilien auftauchen, sind es unterschiedliche, und die meisten haben voll ausgebildete Flügel. Es gibt ein paar primitivere flügellose Formen, doch es sind keine überzeugenden Übergangsformen bekannt. 144
Ein Hauptcharakteristikum der Fliegen ist ihre bewundernswerte, verblüffende Flugtechnik. Während der Mensch seine Arme nicht einmal 10 Mal pro Sekunde auf und ab bewegen kann, bringt es eine durchschnittliche Fliege auf 500 Flügelschläge pro Sekunde, und sie bewegt beide Flügel simultan. Die kleinste Asymmetrie beim Flügelschlag ließe die Fliege die Balance verlieren, doch dies geschieht nie.
In seinem Beitrag "Der mechanische Aufbau von Fliegenflügeln" führt Wootton weiter aus:
Je besser wir die Funktion der Insektenflügel verstehen, umso feiner und wunderbarer erscheinen sie uns... Bauteile werden traditionell so ausgelegt, dass sie einer möglichst geringen Deformation unterliegen, Mechanismen werden so ausgelegt, dass sie Bestandteile in vorhersehbarer Weise bewegen. Insektenflügel kombinieren beides miteinander, sie verfügen über hochelastische Komponenten, die auf die eleganteste Art und Weise zusammengefügt sind, um erforderliche Deformationen unter entsprechender Krafteinwirkung zu ermöglichen und somit das Medium Luft optimal nutzen zu können. Es gibt fast keine Parallelen dazu in der Technik.145
Natürlich kann das plötzliche Auftauchen von Lebewesen mit solch perfektem Design nicht evolutionistisch erklärt werden. Deswegen sagt Pierre-Paul Grassé: "Was den Ursprung der Insekten angeht, tappen wir in Dunkeln."146 Es ist völlig klar, dass der Ursprung der Insekten in der Schöpfung liegt.
Die Herkunft der Säugetiere
Wie wir bereits zuvor dargelegt haben, schlägt die Evolutionstheorie vor, ein paar imaginäre Kreaturen seien aus dem Wasser gekrochen, hätten sich in Reptilien verwandelt und Vögel hätten sich aus den Reptilien entwickelt. Nach demselben Szenario sind Reptilien nicht nur die Vorfahren der Vögel, sondern auch der Säugetiere. Doch zwischen diesen beiden Klassen gibt es große Unterschiede. Säugetiere sind Warmblüter, sie erzeugen ihre Körperwärme selbst und halten sie auf einem konstanten Niveau, sie sind lebend gebärend, sie säugen ihre Jungen und ihre Körper sind mit Fell oder Haaren bedeckt. Reptilien sind dagegen Kaltblüter, ihre Körpertemperatur passt sich der ihrer Umgebung an, sie legen Eier, sie säugen ihre Jungen nicht, und ihre Körper sind mit Schuppen bedeckt.
Wie soll ein Reptil bei all diesen Unterschieden angefangen haben, seine Körperwärme zu erzeugen, woher soll der notwendige Perspirationsmechanismus gekommen sein, die Temperatur konstant zu halten? Ist es möglich, dass es seine Schuppen durch Fell oder Haare ersetzte und anfing, Milch zu geben? Wenn die Evolutionstheorie die Herkunft der Säugetiere erklären will, muss sie zunächst einmal wissenschaftliche Antworten auf diese Fragen geben.
Doch wenn wir uns evolutionistische Quellen anschauen, finden wir entweder imaginäre und unwissenschaftliche Szenarien oder tiefes Schweigen. Eines dieser Szenarien läuft folgendermaßen:
Einige Reptilienarten, die in kälteren Regionen lebten, begannen eine Methode anzuwenden, ihre Körper warm zu halten. Ihre Schuppen wurden kleiner und spitzer und verwandelten sich in ein Fell. Auch die Fähigkeit des Schwitzens war eine Anpassung, die erfolgte, um die Körpertemperatur regeln zu können, ein Mechanismus, den Körper zu kühlen, wenn nötig durch Verdunstung von Wasser. Zufällig begannen nun einige Junge dieser Tiere den Schweiß der Mutter zwecks Nahrungsaufnahme abzulecken. Deswegen sonderten einige der Schweißdrüsen ein immer reichhaltigeres Sekret ab, das später zu Milch wurde. So hatten diese frühen Säugetiere einen vorteilhafteren Start ins Leben.147
Das obige Zitat ist nichts als ein Hirngespinst. Ein solch phantastisches Szenario wird nicht nur von keinerlei Beweis gestützt, es ist völlig unmöglich. Es ist vollständig irrational, zu behaupten, ein Lebewesen beginne, eine so komplex aufgebaute Nahrung wie Milch zu produzieren, in dem es den Körperschweiß der Mutter ableckt.
Der Grund, warum solche Szenarien erfunden werden, liegt in den immens großen Unterschieden zwischen Reptilien und Säugetieren. Ein Beispiel für die strukturellen Barrieren zwischen Reptilien und Säugetieren ist ihre Kieferanatomie. Säugetierkiefer haben einen Unterkiefer, der nur aus einem einzigen Knochen besteht, der die Zähne trägt. Reptilien dagegen haben drei kleine Unterkieferknochen auf jeder Seite des Unterkiefers. Ein weiterer grundsätzlicher Unterschied besteht darin, dass alle Säuger drei Mittelohrknochen aufweisen, nämlich Hammer, Amboss und Steigbügel. Reptilien haben nur einen einzigen Knochen im Mittelohr. Evolutionisten behaupten, Reptilkiefer und Reptilmittelohr hätten sich schrittweise zum Säugetierkiefer und Säugetierohr entwickelt. Die Frage, wie sich ein Ohr mit einem einzigen Knochen zu einem Ohr mit drei Knochen wandeln kann, und wie der Gehörsinn während dieses Prozesses weiter funktioniert haben soll, wird nie erklärt werden können. Da überrascht es nicht weiter, dass bisher nicht ein einziges Fossil gefunden wurde, dass Reptilien und Säugetiere verbinden würde. Darum war auch der renommierte evolutionistische Wissenschaftsautor Roger Lewin gezwungen, zuzugeben: "Der Übergang zum ersten Säugetier... ist immer noch ein Rätsel."148

Es gibt keinen Unterschied zwischen den dutzenden von Millionen Jahre alten Säugetierfossilien in den naturhistorischen Museen und heute lebenden Säugetieren. Auch diese Fossilien tauchen plötzlich auf, ohne dass es eine Verbindung zu vorher lebenden Arten gegeben hätte.
George Gaylord Simpson, eine der wichtigsten Autoritäten unter den Evolutionisten und einer der Begründer der Theorie des Neo-Darwinismus, kommentiert dieses verwirrende Problem der Evolutionisten wie folgt:
Das erstaunlichste Ereignis in der Geschichte des Lebens auf der Erde ist der Wechsel im Mesozoikum, der Wechsel vom Reptilienzeitalter zum Zeitalter der Säugetiere. Es ist, als ob plötzlich der Vorhang vor einer Bühne niedergegangen war, auf der sämtliche Hauptrollen von Reptilien, speziell von Sauriern in großer Zahl und verwirrender Vielfalt gespielt worden waren, um sich sofort wieder zu heben und dasselbe Bühnenbild zu zeigen, doch diesmal mit völlig neuer Besetzung, in der die Saurier überhaupt nicht mehr vorkamen, andere Reptilien nur noch als Statisten und in der alle Hauptrollen von Säugetieren gespielt wurden, auf die es in den vorhergehenden Akten kaum einen Hinweis gab.149
Als die Säugetiere ihren plötzlichen Auftritt hatten, waren sie bereits sehr verschieden voneinander. Fledermäuse, Pferde, Mäuse und Wale sind Säugetiere und sie alle tauchten in demselben geologischen Zeitalter auf. Eine evolutive Beziehung unter ihnen herstellen zu wollen, ist selbst bei ausgeprägtester Vorstellungskraft nicht möglich. Der Zoologe R. Eric Lombard kommt in einem Artikel des führenden Journals Evolution auf diesen Punkt:
Diejenigen, die nach speziellen Informationen suchen, die bei der Verfassung einer Evolutionsgeschichte für Säugetierkategorien nützlich sein könnten, werden enttäuscht sein.150
Kurz, die Herkunft der Säugetiere - wie auch die anderer Gruppen – stimmt in keiner Weise mit der Evolutionstheorie überein. George Gaylord Simpson gestand diese Tatsache schon vor vielen Jahren zu:
Dies gilt für alle 32 Ordnungen der Säugetiere... Die frühesten und primitivsten bekannten Ordnungen (von Säugetieren) haben bereits die grundlegenden Merkmale ihrer Ordnung und in keinem Fall ist ein approximativer, kontinuierlicher Übergang von einer Ordnung zur anderen bekannt. In den meisten Fällen ist der Bruch so glatt und die Kluft ist so breit, dass der Ursprung einer Ordnung spekulativ und Gegenstand vieler Diskussionen ist... Diese regelmäßige Abwesenheit von Übergangsformen beschränkt sich nicht auf Säugetiere, sondern ist ein nahezu universales Phänomen, das von Paläontologen schon lange bemerkt worden ist. Es gilt für fast alle Tierklassen, Wirbeltiere und Wirbellose... es gilt für die Klassen und für die hauptsächlichen Tierabteilungen, und wie es scheint, auch für die analogen Pflanzenkategorien.151
Der Mythos der Evolution von Pferden
Ein wichtiges Thema bei der Frage nach der Herkunft der Säugetiere ist der Mythos von der Evolution des Pferdes, ein Thema, dem in evolutionistischen Publikationen seit langem breiter Raum gegeben wird. Ein Mythos ist es deswegen, weil es eher auf Einbildungskraft als auf wissenschaftlichen Entdeckungen beruht.
Bis vor kurzem wurde eine eindrucksvolle, angeblich die Evolution des Pferdes zeigende, gleichwohl imaginäre Fossilienreihe als prinzipieller Fossilienbeweis für die Evolutionstheorie vorgeführt. Heute hingegen geben selbst viele Evolutionisten offen zu, dass das Szenario der Pferdeevolution am Ende ist. 1980 wurde ein viertägiges Symposium im Museum für Naturgeschichte in Chicago abgehalten, an dem 150 Evolutionisten teilnahmen. Thema war die Problematik der graduellen Evolutionstheorie. Der Evolutionist Boyce Rensberger bemerkte in seinem Vortrag, dass das Szenario der Pferdeevolution keine Grundlage im Fossilienbestand habe und dass kein Evolutionsprozess beobachtet worden ist, der die graduelle Evolution von Pferden nachweisen könnte:
Das vielzitierte Beispiel der Pferdeevolution, dass eine Reihe gradueller Veränderungen einer fuchsähnlichen Kreatur vorsah, die vor 50 Millionen Jahren begann und mit dem heutigen wesentlich größeren Pferd endete, ist lange als falsch bekannt. Anstatt dass graduelle Veränderungen zu beobachten wären, taucht jede Zwischenform klar erkennbar getrennt von anderen auf, lebt eine Zeit lang unverändert und stirbt dann aus. Übergangsformen sind nicht bekannt.152
Bei seiner besonders aufrichtigen Abhandlung dieses Dilemmas der Evolution von Pferden brachte Rensberger das Problem der Übergangsformen als das größte aller Probleme auf die Tagesordnung.
Der wohlbekannte Paläontologe Colin Patterson, ein Direktor des naturgeschichtlichen Museums in London, in dem im Untergeschoss zu dieser Zeit Darstellungen der "Evolution des Pferdes" öffentlich ausgestellt wurden, sagte folgendes über diese Ausstellung:
Es hat eine Unmenge an Geschichten gegeben, manche phantasiereicher als andere, was denn nun das Wesen dieser Geschichte (des Lebens) wirklich ist. Das berühmteste Beispiel, das immer noch unten ausgestellt ist, sind die vor etwa 50 Jahren angefertigten Darstellungen der Pferdeevolution. Sie wurden in einem Lehrbuch nach dem anderen als die reine Wahrheit präsentiert. Ich denke, das ist bedauerlich, besonders wenn die Leute, die diese Art Geschichten verbreiten, sich womöglich ganz genau im Klaren darüber sind, wie spekulativ das alles ist.153

Die Ausstellung über die Evolution des Pferdes, gezeigt im Londoner Museum für Naturgeschichte. Diese und andere Darstellungen der "Evolution des Pferdes" zeigen eigenständige Arten, die zu verschiedenen Zeiten an verschiedenen Orten gelebt haben, völlig willkürlich hintereinander aufgereiht. In Wahrheit gibt es überhaupt keine wissenschaftlichen Entdeckungen über eine Evolution des Pferdes.
Was ist dann die Grundlage für das Szenario der Evolution des Pferdes? Es wurde entwickelt mit Hilfe irreführender Diagramme, die durch die Aneinanderreihung ausgestorbener Arten, die zu weit auseinanderliegenden Zeiträumen in Indien, Süd-Afrika, Nord-Amerika und Europa lebten, entstanden und die nur übereinstimmten mit der großen Kraft evolutionistischen Vorstellungsvermögens. Mehr als 20 Darstellungen der Pferde-Evolution, die übrigens alle erheblich voneinander abweichen, wurden von verschiedenen Wissenschaftlern vorgestellt. Dadurch wird auch deutlich, dass Evolutionisten sich nicht einigen konnten über diese Familienstammbäume. Das einzig gemeinsame Merkmal dieser Arrangements ist der Glaube, dass ein Tier von der Größe eines Hundes mit dem Namen Eohippus (Hyracotherium), das vor 55 Millionen Jahren im Eozän lebte, der Vorfahr des Pferdes sei. Tatsache aber ist, dass Eohippus fast identisch ist mit Hyrax, einem kleinen hasenähnlichen Tier, dass immer noch in Afrika lebt und mit einem Pferd nun wirklich überhaupt nichts gemein hat.154
Die Widersprüchlichkeit der These der Pferdeevolution wird umso deutlicher, je mehr Fossilien zusammengetragen werden. Fossilien moderner Pferdearten (Equus nevadensis und Equus occidentalis) sind in derselben Erdschicht wie Eohippus entdeckt worden.155 Das weist klar darauf hin, dass das moderne Pferd und sein so genannter Vorfahr zur selben Zeit lebten.
Der evolutionistische Wissenschaftsautor Gordon R. Taylor erklärt diese wenig bekannte Tatsache in seinem Buch The Great Evolution Mystery (Das große Geheimnis der Evolution):
Doch die wahrscheinlich größte Schwäche des Darwinismus ist das Scheitern der Paläontologen, überzeugende Phylogenesen oder Reihenfolgen zu finden, die einen größeren evolutionären Wandel zeigen... Das Pferd wird oft als das einzige vollständig schlüssige Beispiel gezeigt. Tatsache ist aber, dass die Linie von Eohippus zu Equus extrem unregelmäßig verläuft. Es ist beabsichtigt, ein kontinuierliches Wachstum der Körpergröße zu zeigen, tatsächlich aber waren manche der Varianten kleiner als Eohippus und nicht größer. Exemplare von verschiedenen Fundorten können in eine überzeugend wirkende Reihenfolge gebracht werden, doch es gibt keinen Beweis, dass sie wirklich zeitlich in dieser Reihenfolge auftraten.156
All diese Fakten über die als solider Beweis für den Darwinismus präsentierten Darstellungen der Evolution des Pferdes belegen stichhaltig, dass es sich um nichts als phantastische Märchen handelt. Pferde kamen wie alle anderen Spezies auf die Welt, ohne dass sie Vorfahren im Sinne des Evolutionsgedankens gehabt hätten.
Die Herkunft der Fledermäuse
Eines der interessantesten Tiere in der Klasse der Säugetiere ist ohne Zweifel das fliegende Säugetier, die Fledermaus.
 Das Sonarsystem der Fledermäuse ist empfindlicher und effizienter als irgendein bisher vom Menschen konstruiertes Sonarsystem.
Das Sonarsystem der Fledermäuse ist empfindlicher und effizienter als irgendein bisher vom Menschen konstruiertes Sonarsystem.
Ganz oben auf der Liste der besonderen Eigenschaften der Fledermäuse steht ihr Sonarsystem, dass ihnen ermöglicht, in völliger Dunkelheit zu fliegen und die kompliziertesten Manöver auszuführen, ohne irgendetwas sehen zu können. Sie können sogar eine Raupe auf dem Boden eines dunklen Raumes orten und ergreifen.
Das Fledermaus-Sonar arbeitet folgendermaßen: Das Tier sendet ständig eine kontinuierliche Folge von Ultraschall-Tönen aus und macht sich anhand der von den in seiner Nähe befindlichen Objekten zurückgeworfenen Echos ein detailliertes Bild seiner Umgebung. Dies ist ein permanenter Vorgang, der ohne jede Störung abläuft, während es zielsicher durch die Luft fliegt.
Die Erforschung des Fledermaussonars ergab überraschende Resultate. Der Frequenzbereich, den das Tier wahrnehmen kann, ist sehr schmal, es kann also nur ganz bestimmte Töne hören. Dies sollte eigentlich ein Problem sein für die Fledermaus. Ein von einem sich bewegenden Körper zurückgeworfenes Tonsignal ändert seine Frequenz aufgrund des bekannten Doppler-Effekts. Wenn nun der von einer Fledermaus ausgesendete Ton beispielsweise auf eine sich von ihr weg bewegende Fliege trifft, so hat das die Fledermaus erreichende Echo des Tons eine tiefere Frequenz als der Originalton, sodass das Tier dieses Echo nicht wahrnehmen kann. Daher sollte man annehmen dürfen, dass die Fledermaus allergrößte Schwierigkeiten haben müsste, sich bewegende Körper zu orten.
Dies ist jedoch nicht der Fall. Die Fledermaus fängt alle möglichen kleine, sich schnell bewegende Insekten ohne jede Anstrengung. Sie ändert einfach die Frequenz der ausgesendeten Töne, als wüsste sie über den Doppler-Effekt Bescheid. So sendet sie zum Beispiel das höchste ihr zur Verfügung stehende Tonsignal in Richtung einer sich von ihr wegbewegenden Fliege, sodass die Frequenz des zurückgeworfenen Echos nicht unterhalb des für die Fledermaus wahrnehmbaren Grenzwertes liegt.
Wie aber funktioniert dieser Regelmechanismus?
Es gibt zwei Gruppen von Neuronen (Nervenzellen mit der Fähigkeit, elektrochemische Signale zu erzeugen) im Gehirn der Fledermaus, die das Sonarsystem kontrollieren. Die eine empfängt die Ultraschallechos, die andere steuert die Muskulatur, die die auszusendenden Töne erzeugt. Diese Hirnregionen sind miteinander rückgekoppelt. Wenn eine Empfangsfrequenz sich ändert, nimmt die erste Hirnregion dies wahr und informiert die zweite Hirnregion über die Frequenzänderung. Daraufhin regelt die zweite Region die Sendefrequenz entsprechend der veränderten Empfangsfrequenz nach. Auf diese Weise ändert sich die Tonhöhe der ausgesendeten Ultraschallsignale entsprechend der sich verändernden Umgebung und ermöglicht den effizientesten Einsatz dieses Sonarsystems.
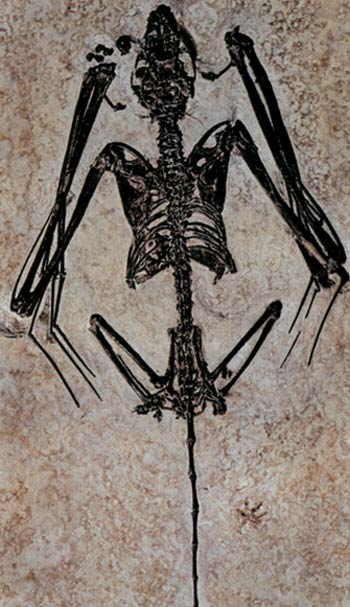
Das älteste bekannte Fledermaus-Fossil, gefunden in Wyoming/USA. Obwohl 50 Millionen Jahre alt, ist keinerlei Unterschied zwischen dem Fossil und heute lebenden Fledermäusen zu erkennen.
Man kann unmöglich so blind sein, nicht zu sehen, dass das Sonarsystem der Fledermaus die Theorie der graduellen Evolution durch zufällige Mutationen vollständig ad absurdum führt. Es handelt sich um eine extrem komplexe Struktur, die unmöglich mit Zufallsmutationen erklärt werden kann. Damit das System überhaupt funktionieren kann, müssen alle seine Komponenten als Ganzes perfekt zusammenarbeiten. Es ist absurd zu glauben, ein so hoch integriertes System könne zufällig entstanden sein. Es zeigt nämlich im Gegenteil, dass es makellos erschaffen wurde.
Tatsächlich bestätigt auch der Fossilienbestand, dass Fledermäuse plötzlich auftauchten und zwar schon mit den heutigen komplexen anatomischen Strukturen. In ihrem Buch Bats: A Natural History (Fledermäuse: eine Naturgeschichte), offenbaren die evolutionistischen Paläontologen John E. Hill und James D. Smith diese Tatsache in Form des folgenden Eingeständnisses:
Der Fossilienbestand der Fledermäuse reicht zurück bis ins frühe Eozän ...und ist dokumentiert worden ...auf fünf Kontinenten ...alle Fledermaus-Fossilien waren ganz klar voll entwickelte Fledermäuse, und so werfen sie denn nur wenig Licht auf ihre Abstammung von ihren am Boden lebenden Vorfahren.157
Der evolutionistische Paläontologe L. R. Godfrey sagt zum selben Thema folgendes:
Es gibt einige bemerkenswert gut erhaltene Fledermaus-Fossilien aus dem frühen Tertiär wie Icaronycteris index, doch Icaronycteris sagt uns nichts über die Evolution des Fluges der Fledermäuse, weil er eine perfekt fliegende Fledermaus war.158
Der evolutionistische Wissenschaftler Jeff Hecht gibt dasselbe Problem in einem Artikel im New Scientist von 1998 zu:
Die Herkunft der Fledermäuse ist ein Puzzle gewesen. Selbst die ältesten Fledermausfossilien von vor über 50 Millionen Jahren haben Flügel, die denen der modernen Fledermäuse fast genau ähneln.159
Kurz, die komplexen biologischen Systeme der Fledermäuse können nicht durch Evolution entstanden sein, und der Fossilienbestand zeigt, dass dies auch nicht geschehen ist. Die ersten Fledermäuse der Welt waren genau dieselben wie die von heute. Fledermäuse sind immer Fledermäuse gewesen und nie etwas anderes.
Die Herkunft der Meeressäuger
Wale und Delphine gehören zur Ordnung der Meeressäuger, der Cetacea. Sie werden als Säugetiere klassifiziert, weil sie wie an Land lebende Säugetiere lebend gebärend sind und ihre Jungen säugen, Lungen zum Atmen haben und Warmblüter sind. Für Evolutionisten war die Herkunft der Meeressäuger eines der schwierigsten Probleme, dass sie zu erklären hatten. In vielen evolutionistischen Quellen wird behauptet, die Vorfahren der Meeressäuger seien vom Land ins Meer gegangen und hätten sich über einen extrem langen Zeitraum hinweg in Meeressäuger verwandelt. Demnach sind die Meeressäuger dem entgegengesetzten Weg des Übergangs aus dem Wasser an Land gefolgt und haben einen zweiten Evolutionsprozess durchlaufen, indem sie ins Wasser zurückgekehrt sind. Dieser Theorie fehlt der paläontologische Beweis und sie widerspricht sich selbst. So haben Evolutionisten denn auch lange Zeit darüber geschwiegen.
In den 90er Jahren jedoch brach neue Begeisterung bei den Evolutionisten aus, glaubten sie doch, die Herkunft der Meeressäuger durch neue Fossilienfunde aus den 1980ern wie dem Pakicetus und Ambulocetus erklären zu können. Diese ausgestorbenen, nachweislich vierbeinigen Landsäugetiere sollten angeblich die Vorfahren der Wale sein, und so zögerten viele evolutionistische Quellen nicht, sie "laufende Wale" zu nennen. Tatsächlich bedeutet der volle Name Ambulocetus natans "laufender und schwimmender Wal". Die übliche Propaganda evolutionistischer Indoktrination verbreitete die Geschichte weiter. National Geographic verkündete im November 2001 das komplette evolutionistische Szenario über die "Evolution der Wale". Doch wie immer basierte dieses Szenario auf evolutionistischer Voreingenommenheit und nicht auf wissenschaftlich bewiesenen Tatsachen.
Der Mythos des laufenden Wals
Fossilienreste des ausgestorbenen Säugetiers Pakicetus inachus tauchten das erste Mal 1983 auf. P. D. Gingerich und seine Assistenten, die das Fossil fanden, zögerten nicht, sofort zu behaupten, es handele um einen "primitiven Wal", obwohl sie tatsächlich nur einen Schädel gefunden hatten.
Das Fossil hat jedoch mit einem Wal absolut nichts zu tun. Sein Skelett erwies sich als das eines Vierbeiners, ähnlich dem des gemeinen Wolfs. Es wurde in einer eisenerzreichen Region gefunden, die voller Fossilien von Landbewohnern war, es fanden sich Schnecken, Schildkröten und Krokodile. Die Region war also terrestrisch gewesen, nicht aquatisch.
Weshalb aber wurde dann ein vierbeiniger Landbewohner als primitiver Wal bezeichnet und warum wird er noch heute von Publikationen wie dem National Geographic als solcher präsentiert? Das Magazin gibt die folgende Antwort:

Gerçek Pakicetus
National Geographic'in Pakicetus çizimi
Entstellende Rekonstruktionen des National Geographic
Paläontologen glauben, Pakicetus sei ein vierbeiniges Säugetier gewesen. Die Skelettstruktur (oben), publiziert im Magazin Nature, zeigt dies deutlich. Die Rekonstruktion des Pakicetus (mitte) von Carl Buell, die darauf basiert, ist demnach realistisch.
National Geographic zog es jedoch vor, ein Bild eines schwimmenden Pakicetus (unten) zu zeigen, um das Tier als "laufenden Wal" darstellen zu können und bei den Lesern diese Vorstellung zu erzwingen. Die Widersprüche in dem Bild sind offensichtlich: Das Tier wurde "schwimmend" dargestellt. Seine Hinterbeine strecken sich rückwärts und sollen so den Eindruck von Flossen vermitteln.
Was bewegt Wissenschaftler dazu, eine Kreatur zu einem Wal zu erklären? Es ist die Gesamtheit vieler subtiler Hinweise - Die Anordnung der Backenzahnkronen, die Biegung eines Mittelohrknochens und die Position der Ohrknochen im Schädel finden sich so bei keinem anderen Landsäuger, sondern ist ein einzigartiges Merkmal der späteren Wale des Eozän.160
Mit anderen Worten, aufgrund von paar Details an Zähnen und Ohrknochen sieht sich das National Geographic in der Lage, diesen vierbeinigen, wolfsähnlichen Landbewohner als laufenden Wal zu beschreiben. Diese Merkmale sind jedoch keineswegs zwingende Beweise, auf denen man eine Verbindung zwischen Pakicetus und dem Wal begründen könnte:Ambulocetus natans: Pençelerine Perde Geçirilen Sahte Balina
Hayali balina evrimi şemasında Pakicetus'tan sonra gelen ikinci fosil canlı, Ambulocetus natans'tır. İlk kez 1994 yılında Science dergisinde yayınlanan bir makaleyle duyurulan bu fosil de, evrimciler tarafından zorlama yöntemiyle "balinalaştırılmak" istenen bir kara canlısıdır.
Ambulocetus natans terimi, Latince ambulate (yürümek), cetus (balina) ve natans (yüzmek) kelimelerinin birleşmesiyle oluşturulmuştur ve "yürüyen ve yüzen balina" anlamına gelir. Canlının yürüdüğü aşikardır, çünkü tüm diğer kara memelileri gibi onun da dört ayağı, hatta bu ayaklara bağlı geniş pençeleri ve arka pençelerinin ucunda toynakları vardır. Ancak canlının bir taraftan da suda yüzdüğü, daha doğrusu yaşamını hem karada hem de suda (amfibi şekilde) sürdürdüğü iddiasının, evrimcilerin ön yargıları dışında, hiçbir dayanağı yoktur.
Bu konuda bilimle hayal gücü arasındaki sınırı görmek için, evrim teorisinin en önde gelen savunucularından biri olan ve Kasım 2001 sayısını "Balinaların Evrimi" propagandasına ayıran National Geographic'in Ambulocetus rekonstrüksiyonuna bir göz atalım. Dergide yayınlanan Ambulocetus çizimi şöyle:
Çizime dikkat ederseniz, bir kara canlısı olan Ambulocetus'u "balinalaştırmak" için yapılmış iki küçük hileyi kolaylıkla fark edebilirsiniz:

• Hayvanın arka bacakları, yürümeye yarayan ayaklar olarak değil de, yüzmeye yarayan yüzgeçler gibi tasvir edilmiş. Oysa gerçekte canlının bacak kemiklerini inceleyen Carroll, bu canlının "kara üzerinde güçlü bir hareket yeteneğine sahip olduğunu" belirtir.163
• Hayvanın ön ayaklarına "palet" görüntüsü verebilmek için perdeler çizilmiştir. Oysa eldeki Ambulocetus fosillerinden böyle bir sonuca varmak mümkün değildir. Gerçekte fosil kayıtlarında, bu gibi yumuşak dokular hemen hiçbir zaman görünmezler. Dolayısıyla canlının iskeleti dışında kalan özellikleri üzerinde yapılan rekonstrüksiyonlar hep spekülatiftir. Bu da evrimcilere geniş bir propaganda malzemesi sunar.
Ambulocetus'un üstteki çizimi üzerinde yapılana benzer evrimci rötüşlarla, her canlıyı, istenen bir başka canlıya benzer gibi göstermek mümkündür. İsterseniz bir maymun iskeletini de, bacaklarını arkaya doğru çizip "yüzgeç" gibi göstermek ve parmakları arasında perdeler çizmek suretiyle, "balinaların atası olan primat" diye sunabilirsiniz.
Ambulocetus fosili üzerinde yapılan bu çizim hilelerinin geçersizliği, yine National Geographic'in aynı sayısında yayınlanan aşağıdaki çizimden anlaşılabilir:
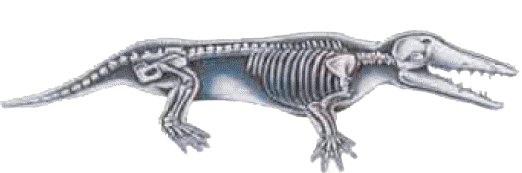
Ambulocetus im National Geographic: Die Hinterbeine des Tieres werden nicht mit Füßen zum Laufen dargestellt, sondern mit Schwimmflossen. Carroll, der die Beinknochen des Tieres untersuchte, sagte jedoch, es habe die Fähigkeit besessen, sich kraftvoll an Land zu bewegen.
National Geographic, canlının iskeletinin resmini yayınlarken, ister istemez rekonstrüksiyon resimde yaptığı "balinalaştırıcı" rötüşlardan geri adım atmak zorunda kalmış. Canlının ayak kemikleri, iskeletin açıkça gösterdiği gibi, onu kara üzerinde taşıyacak yapıda. Ayaklarında ise hayali "perde"lerden iz yok.
Yürüyen Balina Masalının Geçersizliği
Gerçekte ne Pakicetus'un ne de Ambulocetus'un balinalarla bir akrabalıkları bulunduğuna dair hiçbir kanıt yoktur. Bunlar sadece, teorilerine göre deniz memelileri için karada yaşayan bir ata bulmak zorunda olan evrimcilerin, bazı sınırlı benzerliklerden yola çıkarak belirledikleri "ata adayları"dır. Bu canlıların, kendileriyle çok yakın bir jeolojik devirde fosil kayıtlarında ortaya çıkan deniz memelileri ile ilişkileri bulunduğunu gösteren hiçbir kanıt yoktur.
Evrim şemasında Pakicetus ve Ambulocetus'un ardından söz konusu deniz memelilerine geçilmekte ve Procetus, Rodhocetus gibi Archaeocetea (soyu tükenmiş balina) türleri sıralanmaktadır. Söz konusu canlılar gerçekten de suda yaşayan soyu tükenmiş memelilerdir. (Az ileride bunlara da değineceğiz.) Ancak Pakicetus ve Ambulocetus ile bu deniz memelileri arasında çok büyük anatomik farklılıklar vardır. Canlıların fosilleri incelendiğinde, birbirlerine bağlanan "ara form"lar olmadıkları açıkça görülür:
• Dört ayaklı bir kara memelisi olan Ambulocetus'ta omurga, leğen (pelvis) kemiğinde bitmekte ve bu kemiğe bağlı güçlü bacak kemikleri uzanmaktadır. Bu tipik bir kara memelisi anatomisidir. Balinalarda ise, omurga kuyruğa doğru kesintisiz devam eder ve leğen kemiği bulunmaz. Nitekim Ambulocetus'tan 10 milyon yıl kadar sonra yaşadığı düşünülen Basilosaurus aynen bu anatomiye sahiptir. Yani tipik bir balinadır. Tipik bir kara canlısı olan Ambulocetus ile tipik bir balina olan Basilosaurus arasında ise hiçbir "ara form" yoktur.
• Basilosaurus'un ve kaşalotun omurgalarının alt kısmında, omurgadan bağımsız küçük kemikler yer alır. Evrimciler bunların "körelmiş bacaklar" olduğu iddiasındadır. Oysa söz konusu kemikler Basilosaurus'ta "çiftleşme konumunu almaya yardımcı olmakta", kaşalotta ise "üreme organlarına destek olmakta"dır.164 Zaten oldukça önemli bir fonksiyon üstlenmiş olan iskelet parçalarını, bir başka fonksiyonun "körelmiş organı" olarak tanımlamak, evrimci ön yargıdan başka bir şey değildir.
Sonuçta, deniz memelilerinin, kara memelileri ile aralarında bir "ara form" olmadan, özgün yapılarıyla ortaya çıktıkları gerçeği değişmemiştir. Ortada bir evrim zinciri yoktur. Robert Carroll, bu gerçeği istemeden ve evrimci bir dille de olsa, şöyle kabul eder: "Doğrudan balinalara uzanan bir mesonychid çizgisi tanımlamak mümkün değildir."165 Balinalar konusunda ünlü bir uzman olan Rus bilim adamı G. A. Mchedlidze de, bir evrimci olmasına karşın, Pakicetus, Ambulocetus natans ve benzeri dört ayaklı "balina atası adayları"nın bu şekilde tanımlanmasına katılmamakta ve onları tamamen izole bir grup olarak tarif etmektedir.166
Kulak ve Burun Evrimi Hikayeleri
Kara memelileri ile deniz memelileri arasında öne sürülecek bir evrim senaryosunun, bu canlı grupları arasındaki farklı kulak ve burun yapılarına açıklama getirmesi gerekir. Önce kulak yapısını ele alalım. Kara memelileri, biz insanlar gibi, dış dünyadaki sesleri kulak kepçeleri ile toplar, orta kulaktaki kemiklerle güçlendirir ve iç kulakta sinyallere çevirirler. Deniz memelilerinin ise kulakları yoktur. Sesleri alt çenelerindeki özel titreşim algılayıcı duyargalarla duyarlar. Bu iki yapı arasında kademeli bir evrim mümkün değildir. Kendi içinde mükemmel bir duyma sisteminden, tamamen farklı bir yapıya sahip bir başka sisteme kademeli evrimle geçilmesi mümkün değildir. Çünkü ara aşamalar verimli olmayacaktır. Yavaş yavaş kulaklarıyla duyma yeteneğini yitiren, çenesiyle duyma yeteneği ise henüz gelişmemiş bir canlı avantajlı değildir.
Kaldı ki, söz konusu "gelişme"nin nasıl sağlanabileceği sorusu da evrim teorisini çıkmaza sürüklemektedir. Evrimcilerin öne sürdükleri mekanizma mutasyonlardır ve canlılara genetik bilgi ekledikleri hiçbir zaman görülmemiş olan mutasyonlar sonucunda, deniz memelilerinin son derece kompleks algı sistemlerine sahip olduklarını ileri sürmek, akla aykırıdır.
Nitekim fosiller ortada hiçbir evrim olmadığını göstermektedir. Pakicetus ve Ambulocetus'un kulak sistemi, karasal memelilerinki ile aynıdır. Sözde "evrim şeması"nda bu iki kara memelisinin ardından gelen Basilosaurus ise tipik bir balina kulağına sahiptir. Yani dış kulak kepçesiyle değil, çenesine gelen titreşimlerle etrafındaki sesleri algılayan bir canlıdır. Ve Pakicetus ve Ambulocetus'un kulak yapısı ile, Basilosaurus'un kulak yapısı arasında hiçbir "geçiş formu" yoktur.
Benzer bir durum "kayan burun" hikayesi için de geçerlidir. Evrimci kaynaklar, Pakicetus, Rodhocetus ve günümüz gri balinasına ait üç kafatası iskeletini alt alta dizmekte ve bunların bir "evrim süreci" oluşturduklarını ileri sürmektedir. Oysa üç fosilin, özellikle de Rodhocetus ve günümüz balinasının burun yapıları, aynı serinin ara formları olarak kabul edilemeyecek kadar farklıdır.
Dahası nefes deliklerinin burundan enseye doğru "yürümesi", söz konusu canlıların anatomisinde çok ciddi bir "yeniden dizayn" gerektirir ki, bunun rastgele mutasyonlar yoluyla sağlandığına inanmak, hayal kurmaktan başka bir şey değildir.
National Geographic'in Lamarckçı Masalları
Aslında, evrimci camianın büyük bir bölümünün canlıların kökeni hakkında temel bir batıl inanışları vardır ve sorun da bundan kaynaklanmaktadır. Bu batıl inanç, canlılara ihtiyaç duydukları organları, biyokimyasal yapıları veya anatomik özellikleri kazandıran adeta sihirli bir "doğa gücü" olduğu düşüncesidir.
Bunu görmek için, Kasım 2001 tarihli National Geographic'in "Balinaların Evrimi" başlıklı yazısındaki bazı ilginç pasajlara göz atalım:
… Bu civarda bulunan balinaların bazı atalarını gözümün önüne getirmeye çalıştım… Her kuşakta giderek kısalan ve çelimsizleşen arka ayaklarını kullanarak şapıdık şapıdık hareket etmeye çalışıyorlardı… Bir yandan arka bacakları, diğer yandan da gövdelerini destekleyen kalça kemikleri giderek küçülüyordu… Boyun kısaldı, böylece gövdenin ön kısmı, suyu en az dirençle yarıp geçmeyi sağlayan boru biçiminde bir denizaltı gövdesini andırır bir şekle girerken, kollar da dümen biçimini almaya başladı. Dış kulaklara duyulan ihtiyacın azalmasıyla, bazı balinalar sudaki sesleri doğrudan alt çene kemikleriyle algılayıp özel yağ yastıkları üzerinden iç kulağa iletiyorlardı.
Dikkat edilirse, tüm bu anlatımlarda, evrimci mantık örgüsü, sadece canlıların değişen ortama göre değişen ihtiyaçları olduğunu belirtmekte ve bu ihtiyacı başlı başına bir "evrim mekanizması" olarak algılamaktadır: Bu mantığa göre kendisine az ihtiyaç duyulan organlar yok olmakta, ihtiyaç duyulan yeni organlar kendi kendine ortaya çıkmaktadır!
Oysa biyoloji konusunda en temel bilgilere sahip olan bir kimse bile bilir ki, ihtiyaçlarımız organlarımızı kalıtsal olarak şekillendirmez. Bu, Lamarck'ın "kazanılmış özelliklerin sonraki nesillere aktarılması" tezinin çürümesinden bu yana, yani yaklaşık 100 yıldır, bilinen kesin bir gerçektir. Ama evrimci yayınlara bakıldığında, hala Lamarck'ın teorisiyle düşünüyor gibidirler.
Eğer kendilerine itiraz ederseniz, "hayır biz Lamarckçı değiliz, kastımız, çevre şartlarının canlılar üzerinde evrimsel bir baskı oluşturduğu, bu baskı sonucunda uygun canlıların seçildiği ve böylece türün evrimleştiğidir" diyeceklerdir. Ama zaten konunun püf noktası da buradadır: Evrimcilerin "evrimsel baskı" dedikleri şey, canlılara ihtiyaca göre yeni özellikler kazandıramaz. Çünkü bu baskıya cevap vereceğini umdukları iki sözde evrim mekanizması, yani doğal seleksiyon ve mutasyonun canlılara yeni organlar kazandırma özelliği yoktur:
• Doğal seleksiyon, sadece zaten var olan özellikleri seçebilir, yeni bir özellik üretemez.
• Mutasyonlar, canlılara genetik bilgi eklemezler, sadece mevcut genetik bilgiyi tahrip ederler. Genetik bilgi ekleyen, (dolayısıyla yeni bir organ veya biyokimyasal yapı oluşturan) bir mutasyon asla gözlemlenmemiştir.
Bu gerçekler ışığında National Geographic'in üstteki "şapıdık şapıdık hareket eden balinalar" masalına bir kez daha bakarsak, aslında gerçekten de oldukça ilkel bir Lamarckçılık yaptıklarını görürüz. Dikkat edilirse National Geographic yazarı Douglas H. Chadwick, "her kuşakta giderek kısalan ve çelimsizleşen arka ayaklar"dan söz etmektedir. Acaba nasıl olur da "her kuşakta" bir canlı türünde morfolojik değişim, hem de belli bir yöne doğru değişim olabilir? Bunun için; o türün her kuşaktaki kimi temsilcilerinin bacaklarının kısalmasına neden olacak mutasyonlara uğraması; bu mutasyonların canlıya başka hiçbir zarar vermemesi; mutasyona uğrayan bireylerin diğerlerine göre avantajlı olup seçilmesi; bir sonraki kuşakta, ne tesadüfse yine aynı genin aynı noktasının aynı mutasyona uğraması; bunun nesiller boyu hiç değişmeden devam etmesi; tüm bunların tesadüfen kusursuz gerçekleşmesi gerekir.
Eğer National Geographic yazarları buna inanıyorlarsa, "biz sülale olarak uçmayı çok seviyoruz, oğlum da ne tesadüf bir mutasyon geçirdi ve koltuk altlarında kuş tüyünü andıran birkaç küçük yapı belirdi. Torunum da aynı mutasyondan geçecek ve tüyleri biraz artacak, bu nesiller boyu devam edecek ve sonunda sülalemiz kanatlanıp uçacak" diyen bir insana da inanabilirler. İki hikayenin saçmalık düzeyi aynıdır çünkü.
Bu durum, başta belirttiğimiz gerçeği, yani evrimcilerin, canlıların ihtiyaçlarının adeta doğadaki sihirli bir güç tarafından karşılandığına dair batıl inancını ortaya çıkarmaktadır. Gerçekte animist kültürde yer alan "doğaya bilinç atfetme" inancı, ne ilginçtir ki 21. yüzyılda "bilim" kisvesi altında karşımıza çıkmaktadır. Oysa Darwinizm'in ünlü eleştirmenlerinden biri olan Fransa'nın ünlü biyoloğu Paul Pierre Grassé'nin belirttiği gibi, "hayal kurmayı yasaklayan bir kanun yoktur, ama bilim bu işin içine dahil edilmemelidir.167
Fazla sözü edilmeden empoze edilmek istenen bir başka senaryo da, söz konusu canlıların vücut yüzeyleriyle ilgilidir. Karasal canlılar oldukları kabul edilen Pakicetus ve Ambulocetus'un diğer memeliler gibi tüylü bir vücuda sahip oldukları herkesin ortak görüşüdür. Nitekim rekonstrüksiyonlarda her iki canlı sık tüylerle çizilmektedir. Ancak daha sonraki canlılara (yani gerçek deniz memelilerine) geçildiğinde, birden tüyler yok olmaktadır. Bunun evrimsel açıklaması, üstte anlattığımıza benzer Lamarckçı hikayelerden farklı bir şey değildir.
Gerçek ise, söz konusu canlıların her birinin, yaşadıkları ortama göre en uygun biçimlerde yaratılmış olduklarıdır. Bu kusursuz canlıları mutasyonlarla veya daha da basit Lamarckçı hikayelerle açıklamaya çalışmak, akıl dışıdır. Canlılıktaki her özellik gibi, söz konusu canlıların mükemmel sistemleri de bu canlıları Allah'ın yaratmış olduğu gerçeğini gözler önüne sermektedir.
Deniz Memelilerinin Kendi İçindeki Evrimi Senaryosunun Açmazları

Deniz memelileri, yerleştirilmek istendiği hayali evrim şemalarının her birine isyan etmektedirler. Ortaya çıkan sonuç, tarihteki farklı canlı gruplarının, birbirlerinden bağımsız olarak, aniden ortaya çıktıklarıdır. Bu da, tüm canlıların yaratılmış oldukları gerçeğinin bilimsel bir kanıtıdır.
Bu noktaya kadar, deniz memelilerinin kara canlılarından evrimleştiği yönündeki evrimci senaryonun geçersizliğini inceledik. Bilimsel bulgular, evrimcilerin bu senaryonun başlangıcına yerleştirdiği iki kara memelisi (Pakicetus ve Ambulocetus) ile deniz memelileri arasında hiçbir bağ bulunmadığını göstermektedir. Peki senaryonun geri kalan kısmı?
Bu konuda da evrim teorisi yine açmazdadır. Teori, bilimsel sınıflamada Archaeocetea (arkaik, yani eski balinalar) olarak bilinen soyu tükenmiş özgün deniz memelileri ile yaşayan balina ve yunuslar arasında bir akrabalık ilişkisi kurma çabasındadır. Oysa gerçekte konunun uzmanları farklı düşünmektedirler. Evrimci paleontolog Barbara J. Stahl şöyle yazar:
Bu Archaeocetealar'ın kıvrak formdaki vücutları ve kendilerine özgü testere dişleri, bunların muhtemelen herhangi bir günümüz balinasının atası olamayacağını açıkça ortaya koymaktadır. 168
Deniz memelilerinin kökeni konusundaki evrimci senaryo, moleküler biyolojinin bulguları açısından da çıkmaz içindedir. Klasik evrimci senaryo, balinaların iki büyük grubunun, yani dişli balinaların (Odontoceti) ve balenli balinaların (Mysticeti) ortak bir atadan evrimleştiğini varsayar. Ama Brüksel Üniversitesi'nden Michel Milinkovitch yeni bir teoriyle bu görüşe karşı çıkmış, anatomik benzerliğe göre kurulan söz konusu varsayımın moleküler bulgular tarafından çürütüldüğünü şöyle vurgulamıştır:
Cetaceanlar'ın (balinaların) büyük grupları arasındaki evrimsel ilişkiler, morfolojik ve moleküler analizlerin çok farklı sonuçlara varması nedeniyle, daha da problemlidir. Morfolojik ve davranışsal bulgu bütünlerine bakılarak yapılan geleneksel yorumlama, ekolokasyona sahip dişli balinaların (yaklaşık 67 tür) ve filtre sistemiyle beslenen balen balinaların (10 tür) iki ayrı monofilotik (kendi içinde tek kökenden gelen) grup olduğunu varsayar… Öte yandan, DNA üzerinde yapılan filogenetik (evrimsel akrabalık) anazlileri… ve amino asit karşılaştırmaları… uzun zamandır kabul edilen bu sınıflandırmayla çelişmektedir. Dişli balinaların bir grubu, yani sperm balinaları, morfolojik yönden kendilerinden oldukça uzak olan balen balinalarına diğer odontocetlerden (dişli balinalardan) daha yakın gözükmektedirler.169
Kısacası, deniz memelileri, yerleştirilmek istendiği hayali evrim şemalarının her birine isyan etmektedirler.
Deniz memelilerinin kökeni konusundaki evrimci propagandada bir kez daha ortaya çıktığı gibi, ortada gerçek kanıtlara dayanan bir evrim süreci değil, evrim teorisine göre bir şemaya yerleştirilmeye çalışılan, ama bir türlü bu şemaya uygun gelmeyen kanıtlar vardır.
Kanıtların ön yargısız incelenmesiyle ortaya çıkan sonuç ise, tarihteki farklı canlı gruplarının, birbirlerinden bağımsız olarak, aniden ortaya çıktıklarıdır. Bu da, tüm canlıların yaratılmış oldukları gerçeğinin bilimsel bir kanıtıdır.
Memeliler evrim basamaklarının en üst kısmında yer alan canlılar olarak kabul edilirler. Durum bu iken, öncelikle bu canlıların neden deniz ortamına geçtiklerinin açıklanması çok güçtür. Bir sonraki soru ise, bu canlıların deniz ortamına nasıl olup da balıklardan bile daha iyi adapte olduklarıdır. Çünkü katil balinalar, yunuslar gibi memeli ve dolayısıyla akciğerli canlılar, suda solunum yapan balıklardan bile daha mükemmel bir şekilde yaşadıkları ortama uyum göstermektedirler.
Deniz memelilerinin hayali evriminin mutasyon ve doğal seleksiyon aracılığıyla açıklanamayacağı son derece açıktır. GEO dergisinde yayınlanan bir makale, deniz memelilerinden mavi balinanın kökeninden söz ederken, Darwinizm'in bu konudaki çaresizliğini şöyle ifade eder:
Mavi balinalar gibi, denizde yaşayan diğer memeli hayvanların da vücut yapıları ve organları balıklarınkine benzer. Bunların iskeletleri de balıklarınkiyle benzerlik gösterir. Balinalarda bacaklar diyebileceğimiz arka uzuvlar tersine gelişme göstererek güdük kalmıştır. Ancak bu hayvanların şekil değişiklikleri hakkında elde en ufak bir bilgi bile mevcut değildir. Denize geri dönüşün Darwinizm'in iddia ettiği gibi uzun süreli yavaş bir geçişle değil, anlık sıçramalar halinde olduğunu kabul etmek zorundayız. Paleontologlar günümüzde balinanın hangi memeli hayvan türünden geldiği konusunda yeterli bilgiye sahip değildir.170
Karada yaşayan küçük bir memeli hayvanın, evrim süreci sonucunda nasıl olup da 30 metre boyunda 60 ton ağırlığında bir balinaya dönüştüğünü düşünmek gerçekten de çok zordur. Darwinistlerin bu konuda yapabildikleri tek şey, National Geographic dergisinde yayınlanan aşağıdaki anlatımda olduğu gibi, hayal güçlerini zorlayarak senaryo üretmektir:
Balinanın doğuşu, bundan 60 milyon yıl önce, dört ayaklı, kıllı memelilerin yiyecek aramak için denize girmeleriyle başladı. Çağlar geçtikçe, yavaş yavaş değişiklikler oluştu. Arka ayaklar kayboldu, ön ayaklar yüzgeçlere dönüştü, kıllar yok olarak kalın, yumuşak, silgimsi balina derisine yol açtı, burun delikleri başın tepesine hareket etti, kuyruk genişleyerek balinanın fırçamsı kuyruğuna dönüştü ve beden, suyun içinde giderek büyüyüp devleşti.171
Üstte anlatılan kademeli evrim senaryoları, bu senaryoyu yazanlar dahil, hiç kimseyi tatmin etmemektedir. Biz yine de bu kurgunun detaylarına inelim ve ne denli gerçek dışı olduğunu aşama aşama inceleyelim.
Deniz Memelilerinin Özgün Yapıları

Deniz memelileri sadece kendilerine özgün özelliklere sahiptirler. Bunlar, yaşadıkları çevreye en uygun şekilde yaratılmışlardır.
Evrimcilerin deniz memelileri ile ilgili evrim senaryolarının ne kadar imkansız olduğunu gösteren diğer bazı kanıtlar da, bu canlıların son derece özgün yapılarıdır. Solunum için akciğerlerini kullanan memeli bir canlının deniz ortamında geçirmesi gereken adaptasyonlar dikkate alındığında, böyle bir geçiş için "imkansız" kelimesinin bile yetersiz kaldığı görülür. Böyle bir geçişte evrim süreci içinde ara basamaklardan herhangi bir tanesinin bile eksikliği, canlının yaşamasına izin vermeyecek ve evrim sürecini durduracaktır.
Deniz memelilerinin su ortamına geçerken sahip olmaları gereken adaptasyonlar şöyle sıralanabilir:
1- Suyun Korunumu: Deniz memelileri su ortamında yaşamalarına rağmen, su ihtiyaçlarını, balıklar gibi, yani tuzlu sudan faydalanarak gideremezler. Yaşamak için tatlı suya ihtiyaçları vardır. Deniz memelilerinin su kaynakları pek iyi bilinmemesine rağmen, su ihtiyaçlarının büyük kısmını, okyanustaki tuz oranının üçte biri kadar tuz içeren canlıları yiyerek sağladıkları düşünülmektedir. Bu kadar kıt su kaynaklarına sahip deniz memelileri için, suyun azami derecede korunması ve tasarruf edilmesi son derece önemlidir. İşte bu nedenle deniz memelileri, develerde görülen su koruması mekanizmalarına sahiptir. Aynı develer gibi deniz memelileri de terlemez. Böbrekler, üreyi insanlardan çok daha iyi bir şekilde konsantre ederek onlara su kazandırır. Böylece su kaybı en aza indirilmiş olur. Sudan tasarruf en küçük detaylarda bile kendini gösterir. Örneğin anne balina yavrusunu peynir kıvamındaki çok yoğun bir sütle besler. Bu süt insan sütünden on kez daha yağlıdır. Sütün bu derece yağlı olmasının birtakım kimyasal sebepleri vardır. Yağ, yavru tarafından vücuda alındıktan sonra işlenirken yan ürün olarak su açığa çıkar. Böylece anne, en az su kaybıyla yavrusunun su ihtiyacını gidermiş olur.
2- Görme ve Haberleşme: Yunusların ve balinaların gözleri farklı görmelere imkan verecek şekildedir. Suyun altında ve üzerinde aynı mükemmellikte görebilirler. (Oysa başta insan olmak üzere çoğu canlı, ışığın kırılmasındaki farklılıklar nedeniyle, kendi doğal ortamının dışında iyi göremez.) Bir yunus, suyun 6 metre kadar üstüne zıplayabilir ve kendisi için havada tutulmakta olan bir yiyeceği çok büyük bir hassaslıkla alabilir.
Sonuç
Buraya dek incelediğimiz tüm bulgular göstermektedir ki, canlı türleri yeryüzünde her zaman için arkalarında hiçbir evrimsel süreç olmadan, aniden ve kusursuz bir biçimde ortaya çıkmışlardır. Bu durum, evrimci biyolog Douglas Futuyma'nın "canlılar eğer dünya üzerinde eksiksiz ve mükemmel bir biçimde ortaya çıkmışlarsa, o halde üstün bir akıl tarafından yaratılmış olmaları gerekir" derken kabul ettiği gibi,172 canlıların yaratılmış olduklarının çok somut bir ispatıdır.
Evrimciler ise, canlı türlerinin yeryüzünde belirli bir sıra ile ortaya çıkmış olmalarını, evrimleşmiş olduklarının göstergesi gibi yorumlamaya çalışırlar. Oysa canlıların yeryüzündeki ortaya çıkış sıralamaları, ortada hiçbir evrim olmadığına göre, "yaratılışın sıralaması"dır. Fosiller, yeryüzünün, üstün ve kusursuz bir yaratılışla, önce denizlerde sonra da karada yaşayan canlılarla doldurulduğunu ve bütün bunların ardından da insanoğlunun var edildiğini göstermektedir.
DİPNOTLAR
106 John Ostrom, "Bird Flight: How Did It Begin?", American Scientist, Ocak-Şubat 1979, Sayı 67, s. 47.
107 Engin Korur, "Gözlerin ve Kanatların Sırrı", Bilim ve Teknik, Sayı 203, Ekim 1984, s.25.
108 Robert L. Carroll, Patterns and Processes of Vertebrate Evolution, Cambridge University Press, 1997, s. 314.
109 Pat Shipman, "Birds Do It... Did Dinosaurs?", New Scientist, 1 Şubat 1997, s. 28.
110 Pat Shipman, "Birds Do It... Did Dinosaurs?", New Scientist, 1 Şubat 1997, s. 28.
111 Duane T. Gish, Dinosaurs by Design, Master Books, AR, 1996, s. 65-66.
112 Michael Denton, A Theory in Crisis, Adler & Adler, 1986, s. 210-211.
113 Michael Denton, A Theory in Crisis, Adler & Adler, 1986, s. 211-212.
114 Ruben, J. A., T. D. Jones, N. R. Geist and W. J. Hillenius, "Lung Structure And Ventilation in Theropod Dinosaurs and Early Birds", Science 278:1267.
115 Michael J. Denton,. Nature's Destiny, Free Press, New York, 1998, s. 361.
116 Michael J. Denton, Nature's Destiny, Free Press, New York, 1998, s. 361-62.
117 Barbara J. Stahl, Vertebrate History: Problems in Evolution, Dover, 1985, s. 349-350.
118 A. H. Brush, "On the Origin of Feathers", Journal of Evolutionary Biology, vol. 9, 1996, s. 132.
119 A. H. Brush, "On the Origin of Feathers", Journal of Evolutionary Biology, vol. 9, 1996, s. 132.
120 A. H. Brush,"On the Origin of Feathers", Journal of Evolutionary Biology, vol. 9, 1996, s.132.
121 A. H. Brush, "On the Origin of Feathers", Journal of Evolutionary Biology, vol. 9, 1996, s.132.
122 Alan Feduccia, "On Why Dinosaurs Lacked Feathers", The Beginning of Birds, Eichstatt, West Germany: Jura Museum, 1985, s.76.
123 Ernst Mayr, Systematics and The Origin Of Species, Dover, New York, 1964, s. 296.
124 Norman Macbeth, Darwin Retried: An Appeal to Reason, Boston: Gambit, 1971, s. 101.
125 Nature, cilt 382, 1 Ağustos 1996, s. 401.
126 Carl O. Dunbar, Historical Geology, New York: John Wiley and Sons, 1961, s. 310.
127 Robert L. Carroll, Patterns and Processes of Vertebrate Evolution, Cambridge University Press, 1997, s. 280-81.
128 L. D. Martin, J. D. Stewart, K. N. Whetstone, The Auk, cilt 98, 1980, s. 86.
129 L. D. Martin, J. D. Stewart, K. N. Whetstone, The Auk, cilt 98, 1980, s. 86; L. D. Martin "Origins of Higher Groups of Tetrapods", Ithaca, New York: Comstock Publising Association, 1991, s. 485, 540.
130 S. Tarsitano, M. K. Hecht, Zoological Journal of the Linnaean Society, cilt 69, 1985, s. 178; A. D. Walker, Geological Magazine, cilt 177, 1980, s. 595.
131 A. D. Walker, as described in Peter Dodson, "International Archaeopteryx Conference", Journal of Vertebrate Paleontology 5(2):177, Haziran 1985.
132 Richard Hinchliffe, "The Forward March of the Bird-Dinosaurs Halted?", Science, Volume 278, no. 5338, Issue of 24 Oct 1997, s. 596-597.
133 Jonathan Wells, Icons of Evolution, Regnery Publishing, 2000, s, 117.
134 Richard L. Deem "Demise of the 'Birds are Dinosaurs' Theory", http://www.yfiles.com/dinobird2.html
135 Pat Shipman, "Birds do it... Did Dinosaurs?", New Scientist, 1 Şubat 1997, s. 31.
136 "Old Bird", Discover, 21 Mart 1997.
137 "Old Bird", Discover, 21 Mart 1997.
138 Pat Shipman, "Birds Do It... Did Dinosaurs?", s. 28.
139 Ann Gibbons, "Plucking the Feathered Dinosaur", Science, volume 278, no. 5341, Issue of 14 Nov 1997, p. 1229 - 1230.
140 National Geographic, "Feathers for T. Rex?", vol. 196, no. 5, Kasım 1999,
141 Tim Friend, "Dinosaur-bird link smashed in fossil flap", USA Today, 01/25/00.
142 "Open Letter: Smithsonian decries National Geographic's 'editorial propagandizing' of dinosaur-to-bird 'evolution'", http://www.trueorigin.org/birdevoletter.asp
143 M. Kusinitz, Science World, 4 Şubat 1983, s.19.
144 New York Times Press Service, San Diego Union, 29 Mayıs 1983; W. A. Shear, Science, vol. 224, 1984, s. 494.
145 R. J. Wootton, C. P. Ellington, "Biomechanics and the Origin of Insect Flight", Biomechanics in Evolution, ed. J. M. V. Rayner ve R. J. Wootton, Cambridge, Cambridge University Press, 1991, s. 99.
146 J. Robin Wootton, "The Mechanical Design of Insect Wings", Scientific American, cilt 263, Kasım 1990, s. 120.
147 Pierre-P Grassé, Evolution of Living Organisms, New York: Academic Press, 1977, s. 30.
148 George Gamow, Martynas Ycas, Mr. Tompkins Inside Himself, Allen & Unwin, Londra, 1966, s. 149.
149 Roger Lewin, "Bones of Mammals, Ancestors Fleshed Out", Science, cilt 212, 26 Haziran 1981, s. 1492.
150 George Gaylord Simpson, Life Before Man, New York: Time-Life Books, 1972, s. 42.
151 Eric Lombard, "Review of Evolutionary Principles of the Mammalian Middle Ear, Gerald Fleischer", Evolution, cilt 33, Aralık 1979, s. 1230.
152 George G., Simpson, "Tempo and Mode in Evolution", Columbia University Press, New York, 1944, s. 105, 107.
153 Boyce Rensberger, Houston Chronicle, 5 Kasım 1980, Bölüm 4, s. 15.
154 Colin Patterson, Harper's, Şubat 1984, s.60.
155 Francis Hitching, The Neck of the Giraffe: Where Darwin Went Wrong, New York: Ticknor and Fields, 1982, s. 30-31.
156 Francis Hitching, The Neck of the Giraffe, s. 30-31.
157 Gordon Rattray Taylor, The Great Evolution Mystery, London: Sphere Books, 1984, s.230.
158 John E. Hill, James D Smith, Bats: A Natural History, London: British Museum of Natural History, 1984, s. 33.
159 L. R. Godfrey, "Creationism and Gaps in the Fossil Record", Scientists Confront Creationism, W. W. Norton and Company, 1983, s.199
160 Jeff Hecht, "Branching Out", New Scientist, 10 Oct 1998, cilt 160, sayı 2155, s. 14.
161 Robert L. Carroll, Patterns and Process of Vertebrate Evolution, Cambridge University Press, 1998, s. 329.
162 Ashby L. Camp, "The Overselling of Whale Evolution", Creation Matters, a newsletter published by the Creation Research Society, Mayıs/Haziran 1998.
163 Robert L. Carroll, Patterns and Process of Vertebrate Evolution, Cambridge University Press, 1998, s. 333.
164 National Geographic, "Balinaların Evrimi", Kasım 2001, s. 163.
165 Robert L. Carroll, Patterns and Processes of Vertebrate Evolution, Cambridge University Press, 1998, 329.
166 G. A. Mchedlidze, General Features of the Paleobiological Evolution of Cetacea, Rusça'dan Tercüme (Rotterdam: A.A. Balkema), 1986, s. 91.
167 Pierre-P Grassé, Evolution of Living Organisms, New York: Academic Press, 1977, s. 103.
168 B. J. Stahl, Vertebrate History: Problems in Evolution, Dover Publications, Inc., 1985, s. 489.
169 Michel C. Milinkovitch, "Molecular phylogeny of cetaceans prompts revision of morphological transformations", Trends in Ecology and Evolution 10, August 1995, 328-334.
170 Uwe George, "Darwinismus der Irrtum des Jahrhunderts", Geo, Ocak 1984, s. 100-102.
171 Victor B. Scheffer, "Exploring the Lives of Whales", National Geographic, cilt 50, Aralık 1976, s. 752.